 从mRNA到蛋白质.ppt
从mRNA到蛋白质.ppt
- 文档编号:30777529
- 上传时间:2023-08-23
- 格式:PPT
- 页数:181
- 大小:14.95MB
从mRNA到蛋白质.ppt
《从mRNA到蛋白质.ppt》由会员分享,可在线阅读,更多相关《从mRNA到蛋白质.ppt(181页珍藏版)》请在冰豆网上搜索。
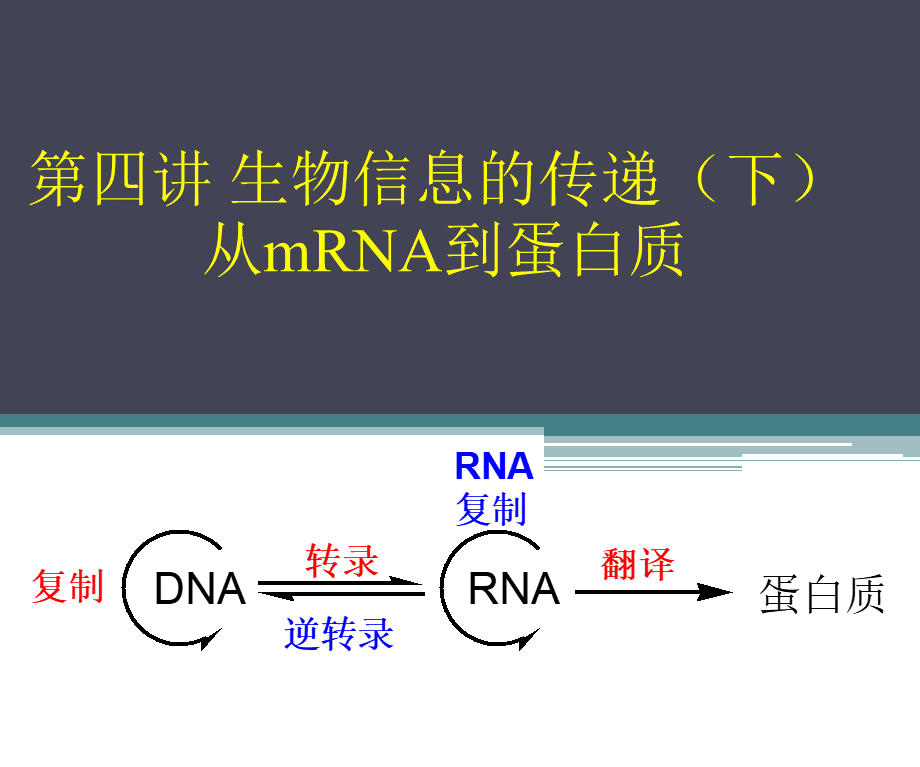
第四讲生物信息的传递(下)从mRNA到蛋白质,蛋白质是生物信息通路上的终产物,一个活细胞在任何发育阶段都需要数千种不同的蛋白质。
活细胞内时刻进行着各种蛋白质的合成、修饰、运转和降解反应。
翻译是指将mRNA链上的核苷酸从一个特定的起始位点开始,按每3个核苷酸代表一个氨基酸的原则,依次合成一条多肽链的过程。
蛋白质的生物合成,蛋白质合成场所:
核糖体;蛋白质合成模板:
mRNA;模板与氨基酸之间的接合体:
tRNA蛋白质合成的原料是:
20种氨基酸,蛋白质合成需要多种蛋白质、酶和其他生物大分子的参与(50%ofthecellsdryweight)蛋白质合成是一个需能反应(80-90%ofthecellsenergy)。
4.1遗传密码三联子,贮存在DNA上的遗传信息通过mRNA传递到蛋白质上,mRNA与蛋白质之间的联系是通过遗传密码的破译来实现的。
遗传密码:
mRNA上每3个核苷酸翻译成多肽链上的一个氨基酸,这3个核苷酸就称为一个密码子(三联子密码)。
mRNA5GCUAGUACAAAACCU3,openingreadingframe(ORF)是指一组连续的含有三联密码子的能够被翻译成为多肽链的RNA序列,它由起始密码子(startcodon)开始,到终止密码子(stopcodon)结束。
4.1.1三联子密码及其破译,若以3个核苷酸代表一个氨基酸,,有43=64种密码子,满足了编码20种氨基酸的需要。
(-)(-)(-),一次删去5-GUAUACGGAU3,读码框不变,用核苷酸的插入或删除实验证明mRNA模板上每三个核苷酸组成一个密码子。
三联子密码的破译,均聚物为模板,Nirenberg把多聚(N)作为模板加入到无细胞体系时发现,新合成的多肽链是:
poly(U)-UUU-多聚苯丙氨酸,poly(C)-CCC-多聚脯氨酸poly(A)-AAA-多聚赖氨酸,poly(G)-成复杂的二级结构而不能被翻译,Poly(UG)-poly(Cys-Val):
5UGUGUGUGUGUGUGUGUG3,无论读码从U开始还是从G开始,都只能有UGU(Cys)及GUG(Val)两种,密码子。
随机共聚物为模板,只含A、C的共聚核苷酸作模板:
8种三联子编码的Asn、His、Pro、Gln、Thr、Lys6种氨基酸。
特定序列的共聚物为模板,以多聚(UUC)为模板,UUC苯丙氨、UCU丝氨酸、CUU亮氨酸,幸运的是由于上述五细胞体系中Mg2+浓度很高,不需要起始密码子可以合成多肽,生理Mg2+条件下,没有起始密码子是不能起始合成的。
以人工合成的三核苷酸如UUU、UCU、UGU等为模板,在含核糖体、AA-tRNA的适当离子强度的反应液中保温后通过硝酸纤维素滤膜。
游离的AA-tRNA因相对分子质量小能自由过膜,与14C标记模板对应的AA-tRNA能与核糖体结合,体积超过膜上的微孔而被滞留。
核糖体结合技术,4.1.2遗传密码的性质,1.密码的连续性(commaless)2.密码的简并性(degeneracy)3.密码的普遍性(universality)4.密码的特殊性(specificity)5.密码子与反密码子的相互作用,编码蛋白质氨基酸序列的各个三联体密码连续阅读,密码间既无间断也无交叉。
由一种以上密码子编码同一个氨基酸的现象称为简并(degeneracy),通用遗传密码及相应的氨基酸,除甲硫氨酸(AUG)和色氨酸(UGG)只有一个密码子外,其他氨基酸都有一个以上的密码子:
9种氨基酸有2个密码子,1种氨基酸有3个密码子,5种氨基酸有4个密码子,3种氨基酸有6个密码子。
AUG甲硫氨酸及起始密码子(常见)GUG缬氨酸及起始密码子(罕见)真核生物的起始密码子总是AUG,UAA终止密码子(赭石,Ochre)UAG终止密码子(琥珀,Amber)UGA终止密码子(蛋白石,Opal),起始密码子和终止密码子,密码的普遍性,生物界基本共用同一套遗传密码。
Thestandardcodonsaretrueformostorganisms,butnotforall,密码子的特殊性,tRNA的反密码子在核糖体内是通过碱基的反向配对与mRNA上的密码子相互作用的。
密码子与反密码子的相互作用,Codon5ACG3Anticodon3UGC5,tRNA反密码子摆动配对(a)tRNALeu的摆动配对;(b)tRNAGly的摆动配对,为每个三联密码子翻译成氨基酸提供了接合体,,为准确无误地将所需氨基酸运送到核糖体上提供了运送载体。
又被称为第二遗传密码。
原核生物中大约有30-45种tRNA,真核细胞中可能存在50种tRNA。
tRNA的3端通过切割、修整,再加上CCA而成;5端由切割产生。
tRNA,tRNA一级结构(PrimaryStructure)tRNA二级结构(SecondaryStructure)tRNA三级结构(TertiaryStructure)tRNA的功能,长度:
60-95nt(commonly76),残基:
15个invariant(恒定)和8个semi-invariant(半恒定).invariant和semi-variant核苷的位置在二级结构和三级结构中起着重要的作用。
含有修饰碱基(Modifiedbases):
有时一个tRNA分子的20%的碱基是经过修饰的。
3端都以CCA-OH结束,tRNA中所有4种碱基都能被修饰,不同tRNA在结构上存在大量的共性,由小片段碱基互补配对形成三叶草形分子结构:
4条手臂(armorstem)3个环(loop),tRNA二级结构(secondarystructure),Tloop,DloopAnticodonloop,tRNA三叶草形二级结构,受体臂(acceptorarm)由链两端序列配对形成的杆状结构和3端未配对的34个碱基所组成。
其3端的最后3个碱基序列永远是CCA,最后一个碱基的3或2自由羟基(OH)可以被氨酰化。
Aminoacidacceptorstem,D环(Darm)的茎区长度常为4bp,也称二氢尿嘧啶环。
负责和氨酰-tRNA合成酶结合,D-armandD-loop,D臂是根据它含有二氢尿嘧啶(dihydrouracil)命名的。
Anticodonloop,反密码子臂是根据位于套索中央的三联反密码子命名的。
由5bp的臂和7个核苷的环组成。
在环中有与密码子互补的由3个核苷组成的反密码子。
它负责对密码子的识别与配对。
VariablearmandT-arm,TC臂是根据3个核苷酸命名的,其中表示拟尿嘧啶;由5bp臂和含有GTC的环组成。
此臂负责和核糖体上的rRNA识别结合;,可变臂(多余臂)是由3到21个核苷组成,可能会形成多达7bp的臂。
tRNA的稀有碱基含量非常丰富,约有70余种。
每个tRNA分子至少含有2个稀有碱基,最多有19个,多数分布在非配对区,特别是在反密码子3端邻近部位出现的频率最高,且多为嘌呤核苷酸。
对于维持反密码子的稳定性及密码子、反密码子间的配对很重要。
在L形三级结构中:
受体臂顶端的碱基位于“L”的一个端点,反密码子臂的套索状结构生成了“L”的另一个端点。
这个结构形式满足了蛋白质合成过程中对tRNA的各种要求而成为tRNA的通式。
tRNA三级结构主要由在二级结构中未配对碱基间形成的9个氢键(三级氢键)而引发的。
大部分恒定或半恒定核苷酸都参与三,级氢键的形成。
tRNA三级结构的形成,模板mRNA只能识别特异的tRNA,而不是氨基酸:
只有tRNA上的反密码子能与mRNA上的密码子相互识别并配对;,氨基酸本身不能识别密码子,只有结合到tRNA上生成AA-tRNA,才能被带到mRNA-核糖体复合物上,插入到正在合成的多肽链的适当位置上。
1.起始tRNA和延伸tRNA2.同工tRNA3.校正tRNA,tRNA的种类,真核生物起始tRNA携带甲硫氨酸(Met),,原核生物起始tRNA携带甲酰甲硫氨酸(fMet),原核生物中Met-tRNAfMet必须首先甲酰化生成fMet-tRNAfMet才能参与蛋白质的生物合成。
1.起始tRNA和延伸tRNA,同工tRNA:
代表相同氨基酸的不同tRNA。
2.同工tRNA,在一个同工tRNA组内,所有tRNA均专一于相同的氨酰-tRNA合成酶。
同工tRNA既要有不同的反密码子以识别该氨基酸的各种同义密码,又要有某种结构上的共同性,能被AA-tRNA合成酶识别。
校正tRNA通过改变反密码子区校正无,义突变和错义突变。
3.校正tRNA,无义突变:
在某个氨基酸的密码子变成终止密码子(UAG、UGA、UAA),使蛋白质合成提前终止,合成无功能的或无意义的多肽,错义突变:
某个核甘酸的变化使一种氨基酸的密码子变为另一种氨基酸的密码子。
无义突变,错义抑制,AA-tRNA合成酶是一类催化氨基酸与tRNA结合的特异性酶:
AA+tRNA+ATPAA-tRNA+AMP+PPi,氨酰-tRNA合成酶,aminonacyl-tRNAsynthetase(ARS),包括两步反应:
第一步:
是氨基酸活化生成酶-氨酰-腺苷酸。
AA+ATP+酶(E)E-AA-AMP+PPi,Aminoacylation(腺苷酰化)oftRNAs,第二步:
是氨酰基转移tRNA3末端腺苷残基的2或3-羟基上,生成氨酰-tRNA。
E-AA-AMP+tRNAAA-tRNA+E+AMP,蛋白质合成的真实性主要决定于AA-tRNA合成酶是否能使氨基酸与对应的tRNA相结合。
AA-tRNA合成酶既要能识别tRNA,又要能识别氨基酸,它对两者都具有高度的专一性。
氨酰-tRNA合成酶利用校读功能来提高精确性。
动力学校对(Kineticprofreading)是基于合成酶对错误氨基酸形状的识别,即使生成氨酰基腺苷酸(AA-AMP)形式,也会将它水解。
构象校对(conformationalproofreading)是在AA-AMP生成后,由于tRNA的进入引起酶构象的改变,使得已生成的AA-AMP不符合进一步连接成AAtRNA的要求,从而水解。
化学校对(Chemicalproofreading)是指错误的氨基酸已连接到tRNA之后,被合成酶的识别位点所发现,使之水解。
1.对应于mRNA中密码子AGG的tRNA反密码子三联体是:
AAGTBGCACCCUDTGC2.不能编码氨基酸的密码子是:
AUAGBAUGCUUGDGUG3.下列哪种突变最可能是致命的?
A插入二个核苷酸B胞嘧啶取代尿嘧啶C缺失三个核苷酸D腺嘌呤取代胞嘧啶4.tRNA的作用是()。
A将一个氨基酸连接到另一个氨基酸上B把氨基酸带到mRNA位置上C将mRNA接到核糖体上D增加氨基酸的有效浓度,5.关于同工tRNA,下列说法错误的是()A.有相同的反密码子环,识别相同的密码子B.结合相同的氨基酸,生成AA-tRNAC.由相同的氨酰-tRNA合成酶催化,合成AA-tRNAD.由不同的反密码子环,识别氨基酸的各种同义密码,左,电镜下看到的胰腺细胞粗糙内质网;右,局部放大后的草图。
结合在内质网上的核糖体,核糖体是一个致密的核糖核蛋白颗粒,可解离为两个亚基,每个亚基都含有一个相对分子质量较大的rRNA和许多不同的蛋白质分子。
核糖体的结构,SizecomparisonsshowthattheribosomeislargeenoughtobindtRNAsandmRNA.,可容纳两个tRNA和约40bp长的mRNA。
原核和真核生物核糖体的组成及功能核糖体亚基rRNAsr-蛋白RNA的特异顺序和功能细菌70S50S23S=2904b36种(L1-L31)2.5106D5S=120b含CGAAC和GTCG互补66%RNA30S16S=1542b21种(S1-S21)16SrRNA(CCUCCU)和SD顺序(AGGAGG)互补哺乳动物80S60S28S=4718b49种4.2106D5S=120b60%RNA5.8S=160b40S18S=1874b33种和Capm7G5端结合,Overviewoftheeventsoftranslation/ribosomecycle,Thelargeandthesmallsubunitsundergoneassociationanddissociationduringeachcycleoftranslation,多聚核糖体(polyribosomesorpolysomes)现象,一个mRNA上连接多个核糖体叫做多聚核糖体,多聚核糖体的意义在于可以同时启动多个翻译进程,能在短时间内能合成较多的肽链,提高翻译的效率。
电镜下的多聚核糖体(polysomes),原核和真核中都存在多聚核糖体现象,在原核细胞中分离的多聚核糖体常与DNA结合在一起,说明RNA尚在转录过程中,因为原核细胞中的转录和翻译是同时进行的,5SrRNA是核糖体大亚基的一个组份,原核生物和真核生物都有5SrRNA,而且结构相似。
真核生物的5SrRNA基因与其它三种rRNA基因不在同一条染色体上,它是由核仁以外的染色体基因转录的,然后运输到核仁内参与核糖体的装配。
5SrRNA基因的数量比45SrRNA转录单位多,人的5SrRNA基因有500个拷贝,并且在染色体上串连排列。
非洲爪蟾的5SrRNA基因的一个重复单位含有一个5SrRNA基因、一个不转录的假基因(101bp的5SrRNA基因的片段),每个重复单位间被不转录的间隔序列隔开,间隔序列的长度变化不定,最长达400bp;5SrRNA基因转录的速度很快,其结果产生过量的5SrRNA,有些最后要被降解掉。
5SrRNA基因是由RNA聚合酶转录的,原初转录物的5端与成熟的5SrRNA的5端完全相同。
在某些生物中,3端通常含有多余的核苷酸,在加工时要被切除。
所以,5SrRNA只需要进行简单的加工,或者根本不需要进行加工。
核糖体小亚基负责对模板mRNA进行序列特异性识别,,大亚基负责携带氨基酸及tRNA的功能,肽键的形成、AA-tRNA、肽基-tRNA的结合等主要在大亚基上。
核糖体的功能,A位点(aminoacylsite),新到来的氨酰-tRNA的结合位点;P位点(peptidylsite),肽酰-tRNA结合位点;E位点(Exitsite),释放去氨酰-tRNA的位点。
只有fMet-tRNAfMet能与第一个P位点相结合,其它所有tRNA都必须通过A位点到达P位点,再由E位点离开核糖体。
每一个tRNA结合位点都横跨核糖体的两个亚基,位于大、小亚基的交界面。
1.核糖体是的合成场所,其上有3个tRNA结合位点,分别是、。
2.核糖体的主要化学成分是和。
根据其在细胞质中存在的状态,可以将其分为和两种类型。
3.细胞中合成蛋白质的场所是()A.溶酶体B.滑面内质网C.细胞核D.核糖体E.细胞质4.真核细胞质中核糖体的大小亚基分别为60S和40S,其完整的核糖体颗粒为()A100SB80SC70SD120SE90S,5组成核糖体的核糖核酸为AmRNABtRNACrRNADsRNAE以上都不是6.真核细胞中由RNA聚合酶负责的核仁rDNA编码的rRNA包括()A.5SrRNA5.8SrRNA18SrRNAB.5SrRNA5.8SrRNA28SrRNAC.5SrRNA18SrRNA28SrRNAD.5.8SrRNA18SrRNA28SrRNAE.以上都不正确,7.核糖体的组装()A.在细胞核任何位置组装成完整核糖体B.在核仁中组装成完整核糖体C.在核仁中分别组装核糖体的亚单位然后在细胞质中组装成完整的核糖体D.完全在细胞质中组装E.有时在细胞核中组装,有时在细胞质中组装8.多聚核糖体是指()A.细胞中有两个以上的核糖体集中成一团B.一条mRNA串连多个核糖体的结构组合C.细胞中两个以上的核糖体聚集成簇状或菊花状结构D.rRNA的聚合体E.附着在内质网上的核糖体,4.4蛋白质的生物合成,氨基酸活化肽链的起始肽链的延伸肽链的终止新合成多肽链的折叠和加工,蛋白质合成各阶段的主要成分简表,1.氨基酸的活化,20种氨基酸,20种氨酰-tRNA合成酶20种以上的tRNA,ATP,Mg2+,氨基酸必须在氨酰tRNA合成酶的作用下生成活化氨基酸AA-tRNA,*同一氨酰-tRNA合成酶具有把相同氨基酸加到多个带有不同反义密码子的同工tRNA分子上的功能。
真核生物起始tRNA是Met-tRNAiMet,原核生物起始tRNA是fMet-tRNAfMet。
tRNA与相应氨基酸的结合是蛋白质合成中的关键步骤,可确保多肽合成的准确性。
蛋白质合成的起始是指:
在模板mRNA编码区5端形成核糖体-mRNA-起始tRNA复合物,并将(甲酰)甲硫氨酸放入核糖体P位点。
2.翻译的起始,原核生物(细菌)为例:
所需成分:
30S小亚基、50S大亚基、模板mRNA、fMet-tRNAfMet、GTP、Mg2+翻译起始因子:
IF-1、IF-2、IF-3、,IF-3,IF-1,翻译起始(翻译起始复合物形成)又可被分成3步:
(P130)1.核蛋白体大小亚基分离,2、30S小亚基通过SD序列与mRNA模板相结合。
IF-3,IF-1,S-D序列,30S亚基具有专一性的识别和选择mRNA起始位点的性质,IF3协助该亚基完成这种选择。
几乎所有原核生物mRNA上都有一个5-AGGAGGU-3序列(SD序列),这个富嘌呤区与30S亚基上16SrRNA3末端的富嘧啶区5-GAUCACCUCCUUA-3相互补。
IF-3,IF-1,3.在IF-2和GTP的帮助下,fMet-tRNAfMet进入小亚基的P位,tRNA上的反密码子与mRNA上的起始密码子配对。
IF-3,IF-1,IF-2,GTP,GDP,Pi,4、GTP水解,释放翻译起始因子,带有tRNA、mRNA的小亚基复合物与50S大亚基结合。
IF-3,IF-1,IF-2,-GTP,GDP,Pi,真核生物翻译起始的特点,核糖体较大,为;起始因子比较多;mRNA5端具有m7Gppp帽子结构Met-tRNAiMetmRNA的5端帽子结构和3端polyA都参与形成翻译起始复合物;40S亚基对mRNA起始密码子的识别经过扫描(Scanning)。
The5endofeukaryoticmRNAiscapped,Kozaksequence真核mRNA通过帽子结合蛋白(eIF-4E能专一地识别帽子结构),起始反应帽子在mRNA与40S小亚基结合过程中起稳定作用带帽子的mRNA5端与18SrRNA的3端序列之间存在不同于SD序列的碱基配对型相互作用,kozak序列是存在于真核生物mRNA的一段序列,其在翻译的起始中有重要作用。
核糖体能够识别mRNA上的kozak序列,并把它作为翻译起始位点。
但是这段序列并不是ribosomalbindingsite(RBS)核糖体结合位点,RBS是帽子结构。
Kozak研究过起始密码子ATG周边碱基定点突变后对转录和翻译所造成的影响,并总结出在真核生物中,起始密码子两端序列为:
G/N-C/N-C/N-ANNATGG,如GCCACCATGG、GCCATGATGG时,转录和翻译效率最高,特别是-3位的A对翻译效率非常重要。
该序列被后人称为Kozak序列,并被应用于表达载体的构建中。
Kozak的“扫描模型”,真核生物核糖体从mRNA的5端(帽子)向包括AUG起始密码子的核糖体结合位点滑动,真核生物翻译起始复合物形成(区别原核生物),原核生物中30S小亚基首先与mRNA模板相结合,再与fMet-tRNAfMet结合,最后与50S大亚基结合。
而在真核生物中,40S小亚基首先与Met-tRNAiMet相结合,再与模板mRNA结合,最后与60S大亚基结合生成80SmRNAMet-tRNAMet起始复合物。
真核生物翻译起始复合物形成过程,生成起始复合物,第一个氨基酸(fMet/Met-tRNA)与核糖体结合以后,肽链开始伸长。
按照mRNA模板密码子的排列,氨基酸通过新生肽键的方式被有序地结合上去。
肽链延伸中的每个循环都包括:
AA-tRNA与核糖体结合;肽键的生成;移位。
肽链的延伸需要,功能核糖体AAtRNA延伸因子(EF-Tu;EF-Ts;EF-G),GTP,Mg2+肽基转移酶,1)后续AA-tRNA与核糖体结合,由于EF-Tu只能与fMet-tRNA以外的其他AA-tRNA起反应,所以起始tRNA不会被结合到A位上,,肽链中间不会出现甲酰甲硫氨酸?
2)肽键的生成,在核糖体mRNAAA-tRNA复合物中,AA-tRNA占据A位,fMet-tRNAfMet占据P位。
生长肽链的C端与P位的tRNA分离,与新的氨基酸之间形成肽键。
构象的变化导致大亚基的移动,使两个tRNA的N端移到大亚基的E和P位,而在小亚基中它们仍位于P和A位。
核糖体通过EF-G介导的GTP水解所提供的能量向mRNA模板3末端移动一个密码子,使两个tRNA完全进入E位和P位(去氨酰tRNA被挤入E位;肽酰-tRNA进入P位),mRNA上的第三位密码子对应于A位准备开始新一轮肽链延伸。
3)移位,延伸因子EF-G有转位酶(translocase)活性,可结合并水解一分子GTP,促进核糖体向mRNA的3方向移动一个密码子的位置,使下一个密码子对准A位点,空载tRNA在E位点脱落。
肽链延伸是由许多个这样的反应组成的:
原核生物中每次反应共需3个延伸因子,EF-Tu、EF-Ts及EF-G,,真核生物细胞需EF-1及EF-2每次循环消耗2个GTP,向生长中的肽链加上一个氨基酸。
当终止密码子UAA、UAG或UGA出现在核糖体的A位时,没有相应的AA-tRNA能与之结合.释放因子能识别这些密码子并与之结合,水解P位上多肽链与tRNA之间的二酯键,释放新生的肽链和tRNA.,核糖体大、小亚基解体,蛋白质合成结束。
释放因子RF具有GTP酶活性,它催化GTP水解,使肽链与核糖体解离。
GTP,mRNA上的终止密码子释放因子,释放因子(终止因子),细菌细胞:
RF1(I类)能识别UAG和UAA,RF2(I类)识别UGA和UAA。
一旦RF与终止密码相结合,它们就能诱导肽基转移酶把一个水分子而不是氨基酸加到延伸中的肽链上。
RF3(II类)与核糖体的解体有关。
真核细胞:
eRF1(I类)能识别三个终止密码子,eRF3(II类),蛋白质合成的能量消耗,每生成一个肽键消耗四个高能键:
1.氨基酸的“活化”消耗2个高能键2.氨酰-tRNA的“进位”消耗一个高能键3.肽酰-tRNA的移位消耗一个高能键合成一个100个氨基酸残基的多肽要消耗多少个高能键?
Termination,Elongation,OverviewoftheeventsoftranslationInitiation,新生的多肽链大多数没有功能,必须经过加工修饰才能转变为活性蛋白质。
蛋白质前体的加工,蛋白质的前体加工包括:
N端fMet或Met的切除二硫键的形成,特定氨基酸的修饰,切除新生肽链中的非功能片段,左:
新生蛋白质在去掉N端一部分残基后变成有功能的蛋白质右:
某些病毒或细菌可合成无活性的多聚蛋白质,经蛋白酶切,割后成为有功能成熟蛋白。
新生蛋白质经蛋白酶切割后变成有功能的成熟蛋白质,1、N端fMet或Met的切除,无论原核生物还是真核生物,N端的甲硫氨酸往往在多肽链合成完毕前就被切除。
2、二硫键的形成,蛋白质的二硫键是蛋白质合成后通过两个半胱氨酸的氧化作用生成的。
二硫键的正确形成对稳定蛋白质的天然构象具有重要的作用。
3、特定氨基酸


- 配套讲稿:
如PPT文件的首页显示word图标,表示该PPT已包含配套word讲稿。双击word图标可打开word文档。
- 特殊限制:
部分文档作品中含有的国旗、国徽等图片,仅作为作品整体效果示例展示,禁止商用。设计者仅对作品中独创性部分享有著作权。
- 关 键 词:
- mRNA 蛋白质
 冰豆网所有资源均是用户自行上传分享,仅供网友学习交流,未经上传用户书面授权,请勿作他用。
冰豆网所有资源均是用户自行上传分享,仅供网友学习交流,未经上传用户书面授权,请勿作他用。
 党组织、党员联系服务师生工作制度.docx
党组织、党员联系服务师生工作制度.docx
 物流与供应链管理 第七章 供应链综合计划.pptx
物流与供应链管理 第七章 供应链综合计划.pptx
-
民族团结教案doc格式.docx
-
民族团结教案doc格式.docx
-
中国象棋ppt模板.pptx
-
设备质量保证措施.docx
-
电力公司招聘行政能力测试试题及解析.docx
 回弹仪检测多孔砖砖强度计算表.xls
回弹仪检测多孔砖砖强度计算表.xls
-
汽车理论1.3.doc
-
电动汽车充换电站项目商业计划书.doc
-
幼儿园指南试题及答案.doc
-
护理教案模板.doc
-
资料台账表格.xlsx
-
红军长征的历史叙述是怎样形成的.docx
-
冷库工程合同.docx
-
部编版五年级上册语文课堂作业本答案(全).docx
-
《网络综合布线》说课稿.doc
-
材料进场,见证取样及检验制度.docx
-
20章--瓦斯综合利用.doc
-
[原创]STP保温板屋面及外墙保温施工技术方案.doc
-
隧道开挖作业台车计算书.docx
-
《减法的初步认识》教学设计.doc
-
教导处工作计划.docx
-
电机学第五版课后答案汤蕴璆完整版.docx
-
同仁堂案例成功启示.docx
-
经典钣金结构设计规范.doc
-
计算机各种进制转换练习题(附答案).doc
-
市场营销教学案例分析.docx
-
服装厂质量管理制度.docx
-
中职试验教材应用文写作(高教版_王崇国编)电子教案【全】.doc
-
控烟考评奖惩制度、标准、记录表.doc
-
小学生“学宪法-讲宪法”活动知识竞赛题库(含答案).docx
-
霍兰德职业倾向测验量表Word文档下载推荐.docx
-
教学实习总结报告Word格式文档下载.docx
-
驾驶员考试题库4高速公路山区道路桥梁隧道夜间恶劣气象和复杂道路条件下的安全驾驶知识127Word格式文档下载.docx
-
果蔬供应链节点分析报告Word文档下载推荐.docx
-
商业项目开发计划书文档格式.docx
-
国内外旅游产业竞争力研究进展讲解Word文档格式.docx
-
高考语文模拟试题4Word格式.docx
-
回填土工程施工方案Word文档下载推荐.docx
-
届山东省菏泽市高三上学期期末考试语文试题B卷 Word版含答案Word下载.docx
-
教育心理学期末总复习文档格式.docx
-
海口初三年级模拟考试二语文含答案文档格式.docx
-
上海市二年级上册数学口算题卡每日30道文档格式.docx
-
海关总署关于非优惠原产地规则中实质性改变标准的规定新Word下载.docx
-
会知识到教会学习的华丽转身Word下载.docx
-
教育营销Word下载.docx
-
某二十六层建筑建给设计Word文档下载推荐.docx
-
混凝土试验室仪器操作规程Word文件下载.docx
-
机关通讯员工作心得Word文档格式.docx
-
韩国交流出国总结Word文档下载推荐.docx
