 4.从mRNA到蛋白质.ppt
4.从mRNA到蛋白质.ppt
- 文档编号:26266362
- 上传时间:2023-06-17
- 格式:PPT
- 页数:71
- 大小:15.26MB
4.从mRNA到蛋白质.ppt
《4.从mRNA到蛋白质.ppt》由会员分享,可在线阅读,更多相关《4.从mRNA到蛋白质.ppt(71页珍藏版)》请在冰豆网上搜索。
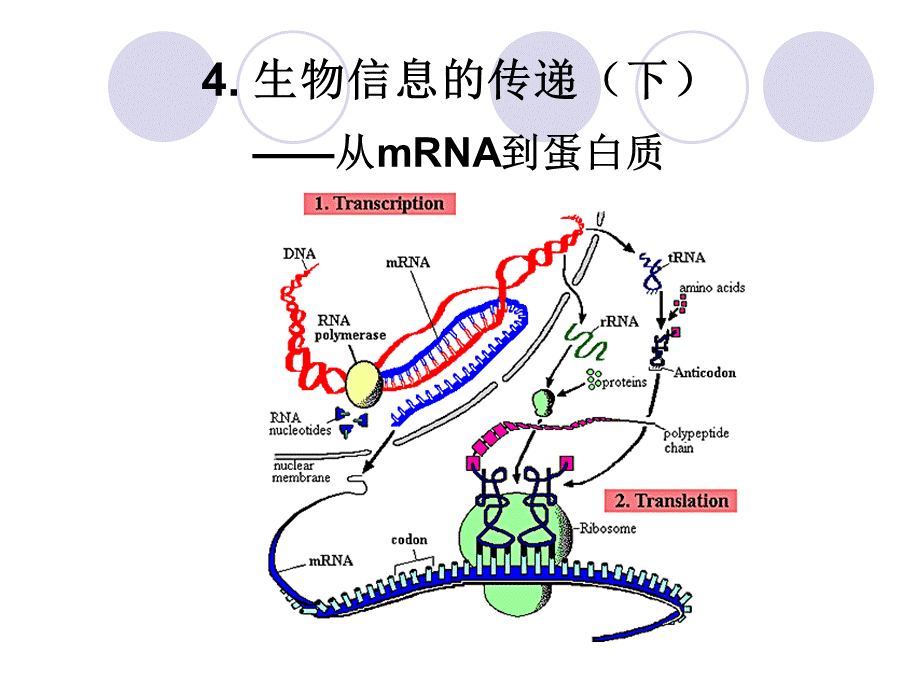
4.生物信息的传递(下)从mRNA到蛋白质,蛋白质是基因表达的产物,生物合成过程包括:
翻译的起始;肽链的延伸;肽链的终止和释放。
核糖体是蛋白质合成的场所,mRNA是蛋白质合成的模板,tRNA是模板与氨基酸之间的接合体。
在合成的各阶段还有许多蛋白质、酶和其他生物大分子参与。
蛋白质合成是一个需能反应,有各种高能化合物参与。
蛋白质合成速度快,大肠杆菌100个氨基酸/5s。
4.生物信息的传递(下)从mRNA到蛋白质,4.1遗传密码三联子4.2tRNA4.3核糖体4.4蛋白质合成的生物学机制4.5蛋白质转运机制,4.1遗传密码三联子,mRNA上每3个核苷酸翻译成蛋白质多肽链上的一个氨基酸,这3个核苷酸称为三联子密码。
翻译时从起始密码子AUG开始,沿着mRNA53连续阅读密码子,至终止密码子为止,生成特定序列的多肽链。
可译框架(openreadingframe,ORF)或可读框。
4.1.1三联子密码及其破译,mRNA中有4种核苷酸,蛋白质中有20种氨基酸,几个核苷酸可以代表20种氨基酸?
在大肠杆菌T4噬菌体中用遗传学方法证实了三联子密码的构想。
对烟草坏死卫星病毒的研究发现,其外壳蛋白亚基由400个氨基酸组成,对应的RNA片段长约1200bp。
3个,用吖啶类试剂处理T4噬菌体(诱导核苷酸插入或丢失),4.1.1三联子密码及其破译,遗传密码的破译(确定代表每种氨基酸的具体密码)得益于体外蛋白质合成体系的建立和核酸人工合成技术的发展。
以均聚物、随机共聚物和特定序列的共聚物为模板指导多肽的合成:
polyUPhe,polyCPro,polyALys,polyGGly;CACHis,CCAPro,AACAsn。
核苷酸结合技术:
人工合成的三核苷酸为模板,与核糖体、AA-tRNA保温,通过硝酸纤维素滤膜,游离的AA-tRNA可通过,与核糖体结合的通不过。
4.1.1三联子密码及其破译,4.1.2遗传密码的性质,密码的连续性翻译从起始密码子开始,一个接一个连续阅读到终止密码子,密码间无间断也无重叠,起始密码子决定了后续密码子的位置。
密码的简并性,4种核苷酸组成64个密码子,61个编码氨基酸,3个是终止密码子,不能与tRNA的反密码子配对,能被终止因子识别,终止肽链的合成。
UAA赭石密码;UAG琥珀密码;UGA蛋白石密码。
61个密码子20个氨基酸,许多氨基酸存在多个密码子。
简并:
由一种以上密码子编码同一个氨基酸的现象,对应于同一氨基酸的密码子称为同义密码子。
密码的简并性,同义密码子其第一、二位核苷酸是相同的,第三位是可变的,这种安排减少了变异的影响。
编码氨基酸的密码子越多,该氨基酸在蛋白质中出现频率越高,Arg例外。
密码的通用性与特殊性,通用性:
遗传密码无论在体内还是体外,对各种生物都是通用的。
特殊性:
支原体中,UGA编码色氨酸;嗜热四膜虫中,UAA编码谷氨酰胺。
密码子与反密码子的相互作用,蛋白质合成中,tRNA的反密码子在核糖体中通过碱基的反向配对与mRNA上的密码子互作。
摆动假说:
密码子与反密码子配对中,前两对遵守碱基配对原则,第三对有自由度,可以“摆动”,使某些tRNA可以识别1个以上密码子。
反密码子第一位为A或C,只识别1种密码子;G或U识别2种;I识别3种。
4.2tRNA,tRNA在蛋白质合成中处于关键地位,为每个三联密码子翻译成氨基酸提供了接合体,还为所需氨基酸运送到核糖体上提供了运送载体,被称为第二遗传密码。
tRNA分子各自的序列不同,但所有的tRNA都具有共同特征:
存在特殊修饰的碱基,3端都以CCA-OH结束,此位点是tRNA与相应的氨基酸结合位点。
tRNA前体的加工,外,tRNA与多种蛋白质和核酸相互识别,决定了在结构上存在共性。
tRNA的三叶草形二级结构,不同的tRNA分子有74-95个核苷酸,长度不同由D臂和多余臂引起。
D臂中存在可变核苷酸位点,包括17:
1、20:
1、20:
2。
根据多余臂的特性将tRNA分为2类:
第一类占75%,含有3-5个核苷酸;第二类含有杆状结构上的5个核苷酸,套索结构上的3-11个核苷酸。
4.2.1tRNA的L形三级结构,tRNA的三级结构呈倒L形折叠,这种结构是二级结构中未配对碱基间形成氢键而引发的,三级结构与AA-tRNA合成酶对tRNA的识别有关。
三级结构中保留了二级结构中由于碱基互补产生的双螺旋杆状结构,又通过分子重排创造了新双螺旋。
tRNA的高级结构满足了蛋白质合成中对其的要求。
4.2.2tRNA的功能,为每个三联密码子翻译成氨基酸提供了接合体,为将所需氨基酸运送到核糖体上提供了运送载体。
氨基酸合成蛋白前通过AA-tRNA合成酶活化,在消耗ATP的情况下结合到tRNA上,生成有蛋白质合成活性的AA-tRNA。
氨基酸本身不能识别密码子,生成AA-tRNA,通过反密码子与mRNA相互识别,才能被带到mRNA-核糖体复合物上,插入到正在合成的多肽链中。
4.2.3tRNA的种类,起始tRNA和延伸tRNA起始tRNA:
特异地识别mRNA上起始密码子的tRNA。
有独特的结构特征,原核生物起始tRNA携带fMet,真核携带Met。
其他tRNA为延伸tRNA。
同工tRNA代表相同氨基酸的tRNA。
同工tRNA组内,所有tRNA均专一于相同的氨酰-tRNA合成酶。
同工tRNA既要有不同的反密码子识别氨基酸的同义密码子,又要有结构上的共同性,能被氨酰-tRNA合成酶识别。
校正tRNA,无义突变:
一个核苷酸的改变可能使代表某个氨基酸的密码子变成终止密码子,蛋白质合成提前终止,合成无功能或无意义的多肽。
校正tRNA可通过改变反密码子区校正无义突变。
错义突变:
由于结构基因中某个核苷酸的变化使一种氨基酸的密码变成另一种氨基酸的密码。
校正tRNA通过反密码子区的改变把正确的氨基酸加到肽链上,合成正常的蛋白质。
校正tRNA,校正tRNA在校正过程中必须与正常的tRNA竞争结合密码子,会影响校正效率,校正效率低于50%。
校正过程还会出现负反应,校正无义突变的同时,还会抑制该基因3端正常的终止密码子,导致通读,蛋白质过多对细胞造成伤害。
一个基因错意突变的校正也可使另一个基因错误翻译。
4.2.4氨酰-tRNA合成酶,一类催化氨基酸与tRNA结合的特异性酶。
反应式:
AA+tRNA+ATPAA-tRNA+AMP+PPi,AA-tRNA合成酶既能识别tRNA,又能识别氨基酸,对两者具有高度的专一性。
使氨基酸与对应的tRNA相结合,保证了蛋白质合成的真实性。
4.3核糖体,生物体内,核糖体像一个沿着mRNA移动的工厂,执行蛋白质合成的功能。
由几十种蛋白质和rRNA组成的亚细胞颗粒。
可以以游离状态存在于细胞内,也可与内质网结合,形成微粒体。
4.3.1核糖体的结构,1)核糖体由大小两个亚基组成原核细胞核糖体70S,由50S和30S两个亚基组成,真核的80S,由60S和40S两个亚基组成。
每个亚基都含有一个相对分子量较大的rRNA和许多蛋白质分子,大分子rRNA能在特定位点与蛋白质结合,从而完成核糖体不同亚基的组装。
2)核糖体蛋白(ribosomalprotein,r-蛋白),核糖体上有多个活性中心,每个中心都由一组核糖体蛋白构成,单个蛋白要在总体结构内才具有催化活性。
E.coli核糖体小亚基由21种r-蛋白组成,用S1-S21表示,大亚基有36种,L1-L36。
真核细胞大亚基含有49种,小亚基33种。
3)核糖体RNA,5SrRNA细菌含有120个(G-)或116个(G+)核苷酸。
两个高度保守区域:
一个含有保守序列CGAAC,是与tRNATC臂上的GTCG互作的部位,是5SrRNA与tRNA相互识别的序列;另一个含有保守序列GCGCCGAAUGGUAGU,与23SrRNA中的序列互补,是5SrRNA与大亚基互作的位点。
3)核糖体RNA,16SrRNA与mRNA、大亚基和tRNA的反密码子直接作用。
存在于小亚基内,1475-1544个核苷酸构成,含有少量修饰碱基。
结构保守,3端保守序列ACCUCCUUA,与mRNA5端翻译起始区富含嘌呤的序列(SD序列)互补;靠近3端还有一段与23SrRNA互补的序列,在大小亚基的结合中起作用。
23SrRNA含有2904个核苷酸,存在于大亚基中,有肽基转移酶的活性。
E.coli第1984-2001存在一段能与tRNAMet互补的片段23SrRNA可能与tRNAMet的结合有关。
靠近5端有一段12个核苷酸的序列与5SrRNA序列互补大亚基上这两种RNA间可能存在互作。
5.8SrRNA真核生物核糖体大亚基特有的rRNA,160个核苷酸,含有修饰碱基。
含有5SrRNA中与tRNA相互识别的保守序列CGAAC可能与原核生物5SrRNA具有相似的功能。
18SrRNA:
存在于真核生物核糖体的小亚基中,酵母的18SrRNA由1789个核苷酸组成,3端与E.coli16SrRNA有同源性。
28SrRNA,长度3890-4500bp,功能未知。
rRNA之间以及与tRNA、mRNA之间的相互关系是建立在序列互补或同源的基础上的。
原核细胞核糖体真核细胞核糖体,4)核糖体有3个tRNA结合位点,A位点:
新进入的氨酰-tRNA的结合位点;P位点:
肽酰-tRNA的结合位点;E位点:
延伸过程中的多肽链转移到氨酰-tRNA上释放tRNA的位点。
tRNA移动顺序是A位P位E位,通过密码子与反密码子间的互作保证反应正向进行。
4)核糖体有3个tRNA结合位点,tRNA的氨基末端定位在大亚基上,另一端的反密码子与结合小亚基的mRNA相互识别,tRNA结合位点横跨核糖体,位于大小亚基的交界面。
4.3.2核糖体的功能,核糖体包括多个活性中心:
mRNA结合部位、A位、P位、肽基转移部位及形成肽键的部位,还有负责肽链延伸的各种延伸因子的结合位点。
小亚基负责对mRNA进行序列特异性识别,如起始部分的识别、密码子与反密码子的互作等,mRNA的结合位点也在小亚基上。
大亚基负责携带氨基酸及tRNA,包括肽键的形成,与AA-tRNA、肽酰-tRNA的结合等。
4.4蛋白质合成的生物学机制,4.4.1氨基酸的活化4.4.2翻译的起始4.4.3肽链的延伸4.4.4肽链的终止4.4.5蛋白质前体的加工4.4.6蛋白质的折叠4.4.7蛋白质合成的抑制剂,4.4.1氨基酸的活化,氨基酸要在氨酰-tRNA合成酶的作用下与tRNA结合,生成活化氨基酸AA-tRNA,才能被运到核糖体中,参与肽链的起始和延伸。
至少存在20种以上具有氨基酸专一性的氨酰-tRNA合成酶,能够识别并通过氨基酸的羧基与tRNA3端腺苷酸核糖基上的3-OH缩水形成二酯键。
4.4.2翻译的起始,细菌中,起始氨基酸是fMet,与核糖体小亚基结合的是N-fMet-tRNAfMet;真核生物中,起始氨基酸是Met,Met-tRNAiMet与小亚基结合起始肽链合成,普通tRNAMet只能进行肽链延伸。
在模板mRNA编码区5形成核糖体-mRNA-起始tRNA复合物,将起始氨基酸放入P位点。
3个步骤:
小亚基先与IF-1、IF-3结合,通过SD序列与模板mRNA结合;在IF-2和GTP帮助下,fMet-tRNAfMet进入小亚基的P位,反密码子与起始密码子配对;带有tRNA、mRNA和翻译起始因子的小亚基复合物与大亚基结合,GTP水解,释放翻译起始因子。
1)原核生物翻译的起始,7种成分:
小亚基;mRNA;fMet-tRNAfMet;翻译起始因子IF-1、IF-2和IF-3;GTP;大亚基;Mg2+。
小亚基通过其16SrRNA的3端与mRNA5端起始密码子上游的SD序列结合,专一性识别和选择mRNA起始位点,IF-3能协助小亚基完成这种选择。
IF-2对于30S起始复合物与50S亚基的连接是必须的,IF-1在70S起始复合物形成后促进IF-2的释放。
2)真核生物翻译的起始,核糖体较大,需要更多的起始因子参与,mRNA的“帽子”和多聚A也参与起始复合物的形成。
40S起始复合物形成中有一种蛋白因子帽子识别蛋白(eIF-4E),专一性识别帽子结构,与mRNA5端结合生成蛋白质-mRNA复合物,通过与eIF-3的亲和力与40S亚基结合。
起始过程需要ATP。
40S亚基还能够识别AUG,用“扫描模型”来解释。
4.4.3肽链的延伸,按照mRNA模板上密码子的排列,每个氨基酸通过缩合反应生成肽键与前一个氨基酸结合,逐渐延长肽链。
肽链延伸有许多循环组成,每加一个氨基酸就是一个循环。
1)后续AA-tRNA与核糖体结合,起始复合物形成后,第二个AA-tRNA在延伸因子EF-Tu及GTP的作用下,生成复合物,结合到A位上。
GTP被水解释放,通过延伸因子EF-Ts再生,形成EF-TuGTP复合物,进入下一轮循环。
模板上的密码子决定了哪种AA-tRNA能被结合到A位上。
EF-Tu只能与起始tRNA以外的AA-tRNA反应。
2)肽键的生成,AA-tRNA2在A位,起始tRNA在P位。
在肽基转移酶的催化下,P位上的氨基酸转移到A位上,与AA-tRNA2上的氨基酸生成肽键二肽酰-tRNA2。
3)移位,核糖体通过EF-G介导的GTP水解提供的能量向向mRNA3端方向移动一个密码子;与第二个密码子结合的二肽酰-tRNA2从A位进入P位,去氨酰-tRNA进入E位,与mRNA上第三位密码子对应的AA-tRNA3进入A位。
4.4.4肽链的终止,终止密码子出现在A位点时,没有AA-tRNA能与之结合,释放因子(RF)能识别并与之结合,水解P位上的多肽链与tRNA之间的二酯键。
释放新生肽链和tRNA,大、小亚基解体,蛋白质合成结束。
释放因子有两类:
类识别终止密码子,类在多肽释放后刺激类从核糖体中解离。
原核与真核生物翻译的比较,4.4.5蛋白质前体的加工,1)N端fMet或Met的切除甲酰基可被脱甲酰化酶水解,Met在多肽合成结束前被切除。
2)二硫键的形成蛋白质合成后通过两个半胱氨酸的氧化作用生成,对稳定蛋白质天然构象有作用。
3)特定氨基酸的修饰包括磷酸化、糖基化、甲基化、乙基化、羟基化和羧基化等。
4)切除新生肽链中的非功能片段多肽类激素和酶的前体都要经过加工切去部分肽段才能成为活性分子。
前胰岛素原蛋白翻译后成熟过程,4.4.6蛋白质的折叠,多肽链的折叠是一个复杂的过程,新生多肽首先折叠成二级结构,然后进一步折叠盘绕成三级结构。
单链多肽蛋白质,三级结构就具备蛋白质的功能;寡聚蛋白质,需要组装成四级结构,才有蛋白活性。
有些蛋白质要在另一些蛋白存在的情况下才能完成折叠过程。
分子伴侣:
序列上没有相关性但有共同功能的保守性蛋白质,在细胞内能帮助其他多肽进行正确的折叠、组装、运转和降解,本身不参与最终产物的形成,如热休克蛋白和伴侣素。
4.4.7蛋白质合成的抑制剂,主要是一些抗生素,如嘌呤霉素(AA-tRNA的结构类似物);氯霉素(阻止mRNA与核糖体结合);四环素(阻止AA-tRNA与核糖体结合);链霉素、新霉素、卡那霉素(干扰AA-tRNA与核糖体结合而产生错读)等。
还有如5-甲基色氨酸、环己亚胺、白喉毒素、干扰素、蓖麻蛋白和其他核糖体灭活蛋白等。
抑制剂用于研究蛋白质的合成机制,也是临床上治疗细菌感染的重要药物。
4.5蛋白质转运机制,细胞各部分都有特定的蛋白质,合成的蛋白要准确地定向运送到细胞的各部分,才能保证生命活动的进行。
4.5.1翻译-转运同步机制4.5.2翻译后转运机制4.5.3核定位蛋白的转运机制4.5.4蛋白质的降解,4.5.1翻译-转运同步机制,信号肽假说认为,蛋白质跨膜转运信号也是由mRNA编码的。
起始密码子后有一段编码疏水性氨基酸的序列信号序列,在核糖体上合成后与膜上特定受体相互作用,产生通道,使这段多肽在合成的同时穿过膜结构。
信号肽的特点:
带有10-15个疏水性氨基酸;N端常有1个或多个带正电荷的氨基酸;C端靠近蛋白酶切割位点处带有多个极性氨基酸,离切割位点最近的氨基酸带有很短的侧链。
蛋白质跨膜转运的信号肽假说及其运输过程,信号识别蛋白,停靠蛋白(DP),4.5.1翻译-转运同步机制,信号肽在蛋白质运输中的作用特点:
完整的信号肽是保证蛋白质转运的必要条件;仅有信号肽还不足以保证蛋白质转运的发生,还要求转运蛋白质在信号序列以外的部分有相应的结构变化;信号序列的切除并不是转运必需的;并非所有的转运蛋白质都有可降解的信号肽。
4.5.2翻译后转运机制,1)线粒体蛋白质跨膜转运通过线粒体膜的蛋白质在转运前大多数以前体形式存在,由成熟蛋白质和位于N端的一段前导肽组成;蛋白质跨线粒体内膜转运是需能过程;蛋白质跨线粒体膜转运时,需要Tom受体复合蛋白和分子伴侣。
2)前导肽的作用和性质,前导肽含20-80个氨基酸,对线粒体蛋白质的识别和转运起关键作用,当前体蛋白跨膜后,其被水解,前体蛋白转变为成熟蛋白,失去跨膜能力。
特性:
带正电荷的碱性氨基酸含量多,分散于不带电荷的氨基酸之间;缺少带负电荷的酸性氨基酸;羟基氨基酸含量高;有形成两亲螺旋结构的能力。
前导肽跨膜转运时先与线粒体外膜上的受体结合,不同部位发挥不同作用。
2)前导肽的作用和性质,3)叶绿体蛋白质跨膜转运,叶绿体定位信号肽有两个部分:
第一部分决定该蛋白质能否进入叶绿体基质;第二部分决定能否进入类囊体。
3)叶绿体蛋白质跨膜转运,特点:
活性蛋白水解酶位于叶绿体基质内,是可溶性的,这是鉴定翻译后转运的指标;叶绿体膜上有识别叶绿体蛋白的受体,能特异地与叶绿体蛋白的前体结合;叶绿体蛋白质前体内可降解序列因植物和蛋白质种类不同而有明显差异。
4.5.3核定位蛋白的转运机制,细胞质中合成的蛋白质通过核孔进入细胞核,核孔是进行双向转运的分子通道。
细胞分裂时,核膜被破坏,分裂完成后,核膜重建,分散在细胞中的核蛋白重新运到核内。
因此,蛋白质中的信号肽核定位序列(NLS)不能被切除,可位于核蛋白的任何部位。
蛋白质向核内运输需要核转运因子、和GTP酶(Ran)参与。
4.5.4蛋白质的降解,大肠杆菌中,蛋白质的降解通过一个依赖于ATP的蛋白酶(Lon)来实现,每切除一个肽键要消耗2个ATP。
真核生物中,蛋白质的降解依赖于泛蛋白,被降解的蛋白在ATP的作用下与泛蛋白相连形成复合体,此过程需要三个降解因子参与,复合体被运送到蛋白降解体系中降解。
蛋白质的半衰期与N-端氨基酸残基的关系,习题,名词1.氨酰tRNA合成酶2.SD序列3.三叶草结构和L结构4.简并性和同义密码子5.起始和终止密码子6.同工tRNA7.起始因子和释放因子8.信号肽9.无义突变和错义突变10.分子伴侣,简答1.遗传密码的性质有哪些?
2.为什么说tRNA是第二遗传密码,有哪些种类的tRNA,各自的功能是什么?
3.原核生物的蛋白质合成可分为哪些阶段?
简述各阶段主要事件。
4.tRNA、mRNA和rRNA在蛋白质合成中的功能各是什么?
5.肽链延伸包括哪些过程?
6.简述原核和真核细胞在翻译过程中的差异。


- 配套讲稿:
如PPT文件的首页显示word图标,表示该PPT已包含配套word讲稿。双击word图标可打开word文档。
- 特殊限制:
部分文档作品中含有的国旗、国徽等图片,仅作为作品整体效果示例展示,禁止商用。设计者仅对作品中独创性部分享有著作权。
- 关 键 词:
- mRNA 蛋白质
 冰豆网所有资源均是用户自行上传分享,仅供网友学习交流,未经上传用户书面授权,请勿作他用。
冰豆网所有资源均是用户自行上传分享,仅供网友学习交流,未经上传用户书面授权,请勿作他用。
 党组织、党员联系服务师生工作制度.docx
党组织、党员联系服务师生工作制度.docx
 物流与供应链管理 第七章 供应链综合计划.pptx
物流与供应链管理 第七章 供应链综合计划.pptx
-
民族团结教案doc格式.docx
-
民族团结教案doc格式.docx
-
中国象棋ppt模板.pptx
-
设备质量保证措施.docx
-
电力公司招聘行政能力测试试题及解析.docx
 回弹仪检测多孔砖砖强度计算表.xls
回弹仪检测多孔砖砖强度计算表.xls
-
汽车理论1.3.doc
-
电动汽车充换电站项目商业计划书.doc
-
幼儿园指南试题及答案.doc
-
护理教案模板.doc
-
资料台账表格.xlsx
-
红军长征的历史叙述是怎样形成的.docx
-
冷库工程合同.docx
-
部编版五年级上册语文课堂作业本答案(全).docx
-
《网络综合布线》说课稿.doc
-
材料进场,见证取样及检验制度.docx
-
20章--瓦斯综合利用.doc
-
[原创]STP保温板屋面及外墙保温施工技术方案.doc
-
隧道开挖作业台车计算书.docx
-
《减法的初步认识》教学设计.doc
-
教导处工作计划.docx
-
电机学第五版课后答案汤蕴璆完整版.docx
-
同仁堂案例成功启示.docx
-
经典钣金结构设计规范.doc
-
计算机各种进制转换练习题(附答案).doc
-
市场营销教学案例分析.docx
-
服装厂质量管理制度.docx
-
中职试验教材应用文写作(高教版_王崇国编)电子教案【全】.doc
-
控烟考评奖惩制度、标准、记录表.doc
-
小学生“学宪法-讲宪法”活动知识竞赛题库(含答案).docx
-
精品县领导干部述职述廉报告.docx
-
江苏省届高三第一次月考 物理.docx
-
九型人格测试题.docx
-
鲁教版小学四年级品德与社会下册教案.docx
-
人教版四年级数学下册小数加减法简便计算专项练习题59.docx
-
年产2亿条塑料编织袋项目可行性研究报告.docx
-
商业计划书研究报告互联网+XX市体育馆建设项目可行性研究报告精品模板.docx
-
推力滚子轴承项目可行性研究报告.docx
-
相城区漕湖清淤取土及湖岸景观工程项目可行性论证报告.docx
-
小学二年级上册《生活生命与安全》教案.docx
-
校园通APP软件设计与实现可行性报告.docx
-
新课标人教版小学四年级上册语文全册教案.docx
-
学年新教材人教版 小学数学五年级上册全册教学案.docx
-
初级轧钢精整工合并卷.docx
-
一建工程经济复习总结.docx
-
教育培训机构创业计划书精编.docx
-
电大经济法律基础电大网络形考13作业.docx
-
中国开关稳压电源供应器行业现状及未来五年投资空间研究报告.docx
-
高三文科综合第一学期期末调研考试.docx
