 免疫学笔记复习知识点总结Word格式.docx
免疫学笔记复习知识点总结Word格式.docx
- 文档编号:21567975
- 上传时间:2023-01-31
- 格式:DOCX
- 页数:14
- 大小:34.58KB
免疫学笔记复习知识点总结Word格式.docx
《免疫学笔记复习知识点总结Word格式.docx》由会员分享,可在线阅读,更多相关《免疫学笔记复习知识点总结Word格式.docx(14页珍藏版)》请在冰豆网上搜索。
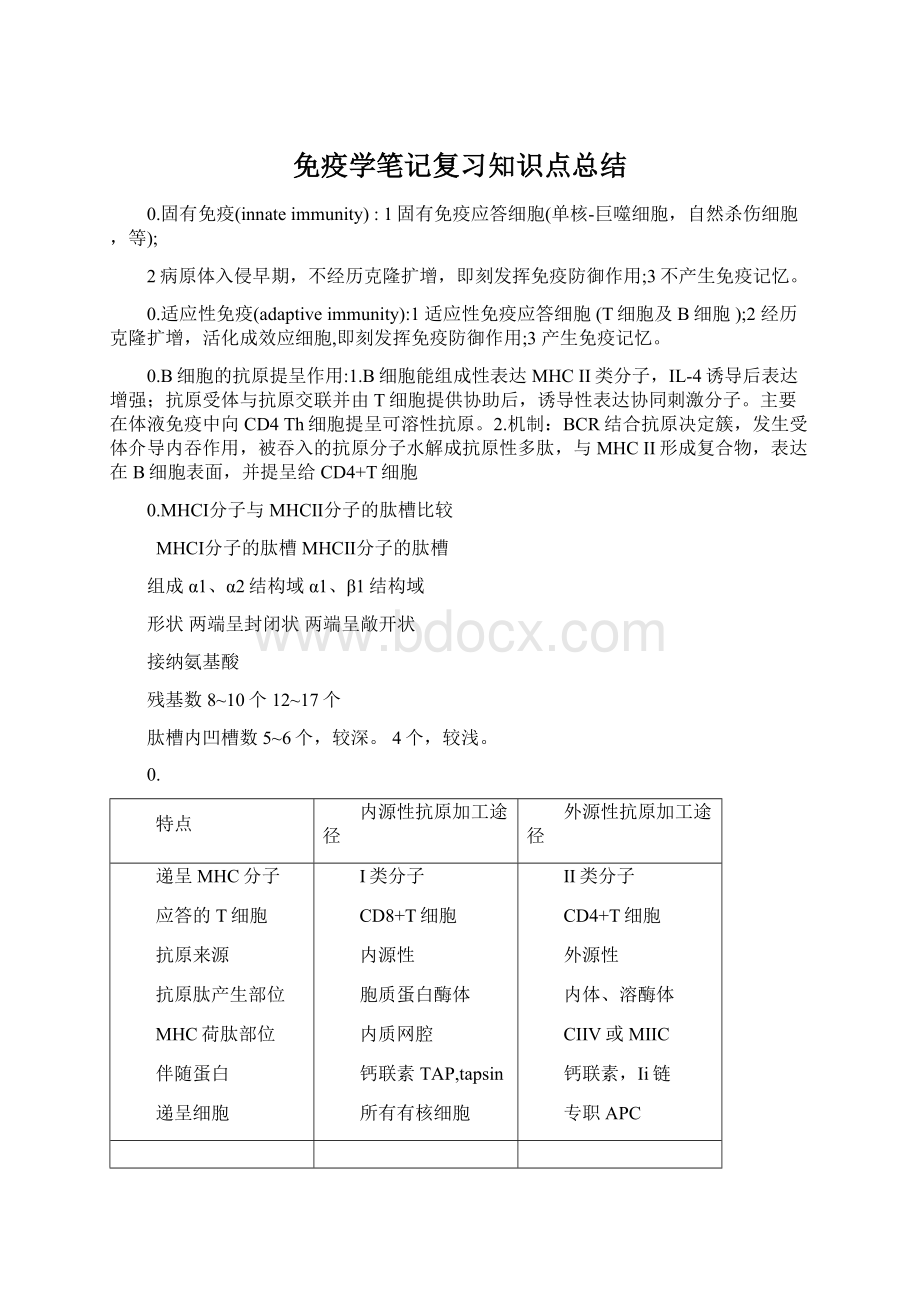
I类分子
CD8+T细胞
内源性
胞质蛋白酶体
内质网腔
钙联素TAP,tapsin
所有有核细胞
II类分子
CD4+T细胞
外源性
内体、溶酶体
CIIV或MIIC
钙联素,Ii链
专职APC
固有免疫应答
适应性免疫应答
主要参与的细胞
黏膜上皮细胞、吞噬细胞、树突状细胞、NK细胞、NKT细胞、gdT细胞、B1细胞
abT细胞、B2细胞、抗原提呈细胞
主要参与的分子
补体、细胞因子、抗菌蛋白、酶类物质
特异性抗体
作用时相
即刻-96小时
96小时后启动
识别受体
膜式识别受体,胚系基因直接编码,较少多样性
特异性抗原识别受体,胚系基因片断发生重排,具有高度多样性
识别特点
直接识别病原体某些共有高度保守的分子结构,具有识别“非己”的能力
T细胞识别APC提呈的抗原肽-MHC分子复合物;
B细胞直接识别抗原表位具有高度特异性
作用特点
不经克隆扩增和分化,迅速产生免疫作用,没有免疫记忆功能
经克隆扩增和分化,成为效应细胞后发挥免疫作用,具有免疫记忆功能
维持时间
维持时间较短
维持时间较长
0淋巴细胞归巢(lymphocytehoming):
成熟淋巴细胞离开中枢免疫器官后,经血液循环趋向性迁移并定居于外周免疫器官或组织的特定区域。
0.淋巴细胞再循环(lymphocyterecirculation):
淋巴细胞在血液、淋巴液、淋巴器官或组织间反复循环的过程。
定居在外周免疫器官(淋巴结)的淋巴细胞,可由输出淋巴管经淋巴干、胸导管或右淋巴导管进入血液循环;
淋巴细胞随血液循环到达外周免疫器官后,可穿越HEV,并重新分布于全身淋巴器官和组织.
0.树突状细胞(Dendriticcells):
摄取、加工并递呈抗原;
参与T细胞亚群的分化;
参与中枢和外周免疫耐受的诱导;
参与调节B细胞功能;
参与固有免疫应答
0.TCR/CD3复合体:
中的两个多态型亚单位(TCRαβ或TCRγδ)主要功能是识别结合MHC分子的抗原,而胞浆区非常短;
CD3分子的主要功能是参与TCR/CD3复合体的装配和稳定以及信号转导.简单的说,也就是APC或B细胞提呈的抗原由TCR-CD3复合体识别,TCR为T细胞表面的抗原识别受体,CD3的作用则是将T细胞识别的抗原信号(第一信号)转导进入T细胞内。
0.T细胞在胸腺发育过程中基因表达的详细分析:
Thegeneticmechanismsthatpromotelineagecommitmentandeliminateautoreactivecellsinthethymusarenotwellunderstood.Tobetterunderstandthisprocess,wehaveidentifiedandquantitatedtranscriptsinthetwomajorthymocytelineagesbyusingserialanalysisofgeneexpression.Approximately25genesdisplayedalmostcompletesegregationtooneortheotherTcelllineage.CommitmenttotheCD4lineagewasmarkedbyup-regulationofgenesassociatedwithincreasedsurvivalandchaperonefunctionfollowedbyexpressionofgenesthatregulatenucleosomeremodelingandTcellreceptorsignaling.DifferentiationwithintheCD8lineage,ontheotherhand,wasmarkedbyup-regulationofgenesthatregulatelymphocytehoming,followedbyquenchingofgenesthatinhibitapoptosis.DefinitionofdifferentialgeneexpressionduringdevelopmentofthetwomajorthymocytelineageswillallowinsightintomechanismsofTcelldevelopmentafterpositiveandnegativeselection.
0.T淋巴细胞
(一)T淋巴细胞在胸腺中的发育
T淋巴细胞简称T细胞。
在胸腺中分化成熟后,进入外周,通过血液和淋巴在全身二级淋巴器官和组织间循环。
T细胞在胸腺内的成熟包括TCR基因重排、阳性选择和阴性选择阶段。
1.TCR基因重排产生功能性TCR基因。
此时T细胞表面同时表达CD4和CD8分子,所以称为双阳性(DP)细胞。
2.阳性选择
DP细胞与胸腺皮质上皮细胞相互作用,凡是其TCR能够识别皮质上皮细胞上自身MHC分子的DP细胞通过选择而存活,否则将在3-4天内凋亡。
所以,阳性选择的结果产生能够识别自身MHC分子的,即受自身MHC分子限制的胸腺细胞。
这种细胞只表达CD4或CD8之中的一种,故称为单阳性细胞,简称SP细胞。
3.阴性选择
通过了阳性选择的SP细胞进入髓质。
在髓质中,凡是其TCR能与DC和Mf上的自身肽-MHC高亲和力结合的细胞,即自身反应性细胞,均发生凋亡。
所以,阴性选择的结果是清除自身反应性T细胞,产生对自身抗原耐受的T细胞。
(二)T细胞表面分子及功能
1.TCR-CD3复合体
1)TCR
TCR是T细胞特有的抗原受体,是T细胞识别抗原的功能性结构。
外周和胸腺中95%的T细胞表达abTCR,少数T细胞表达gdTCR。
a链和b链都是跨膜糖蛋白,两者通过二硫键共价连接。
a和b每条链均包含胞外、跨膜和胞内三部分。
a和b链胞外段都由2个Ig样结构域组成,2条链的胞外段通过折叠后形成的结构与Ig的Fab段非常相似。
a和b链V区中变异程度最大的部位也称为互补决定区(CDRs)。
当T细胞识别MHC-肽时,CDR3与抗原肽相互作用,而CDR1和CDR2则与MHC分子抗原结合部位上的2条a螺旋相互作用。
2)CD3分子
CD3分子与TCR通过非共价结合组成TCR-CD3复合体。
CD3分子是TCR在细胞膜表面稳定表达所必需的,并且转导TCR接受的抗原特异性信号。
CD3是一种由6条跨膜肽链组成的复合体。
有g、d、e、e、x、x与g、d、e、e、x、h二种形式。
CD3分子中每条链的胞内段均含有免疫受体酪氨酸活化基序(ITAM)。
ITAM中的酪氨酸是特异性蛋白激酶催化的目标,它是T细胞活化所必需的。
gdTCR结构与abTCR类似,并且也与CD3分子构成复合体。
2.CD4和CD8分子
1)CD4分子:
CD4分子是一种跨膜糖蛋白,其胞外段含4个Ig样结构域,N端的头2个结构域能与MHCII类分子b链结合,在TCR识别MHCII类-肽的时候,加强CD4+T细胞与APC之间结合的强度,使TCR识别MHCII类-肽复合物的灵敏度提高100倍。
2)CD8分子:
CD8分子也是跨膜糖蛋白,通常以ab异二聚体的形式存在。
每条链的胞外段N末端有一个Ig样的结构域,可与MHCI类分子结合。
3.CD28和CTLA-4(CD152)
CD28是协同刺激分子B7(CD80/CD86)的受体。
当TCR识别APC表面MHC分子-肽复合物时,CD28与APC上的B7分子配接,为T细胞激活提供第二活化信号。
CTLA-4(CD152)也是B7的受体。
只表达在活化T细胞上。
CTLA-4与B7配接后向细胞发送抑制性信号,下调T细胞的功能,阻止活化了的T细胞继续活化,具有重要的免疫调节作用。
4.CD45
CD45又称淋巴细胞共同抗原(leukocytecommonantigen),它是一种蛋白酪氨酸磷酸酶。
其胞内段具有磷酸酶结构域,能使蛋白磷酸激酶Lck和Fyn中的一个酪氨酸去磷酸化而活化,在T细胞活化的启动中起重要作用。
CD45有3种同工型,其中CD45RA表达在初始T细胞表面,CD45RO表达在效应T细胞和记忆T细胞表面。
CD45RA和CD45RO两者均能与CD4和CD8结合,但是CD45RO与CD4和CD8的亲和力高于CD45RA,因此效应细胞对第一信号比初始细胞敏感,不需要第二信号就能对靶细胞或APC上的MHC-肽复合物产生应答。
5.细胞因子受体
T细胞表面表达的细胞因子受体的种类多达十几种。
一个T细胞具体表达的细胞因子受体的种类依细胞的分化阶段、抗原刺激和活化状态等因素而定。
细胞因子受体与自分泌或旁分泌的细胞因子结合,参与细胞的各种活动。
细胞因子受体一般由2个或2个以上的亚单位组成。
细胞因子与相应受体结合后,诱导受体链聚合,激活细胞内信号发送途径(例如激酶级联反应),产生活化的转录因子。
转录因子进入细胞核,与基因的启动子或增强子结合,使有关基因转录而表达相应的功能。
6.CD40L
CD40L表达在活化的Th细胞表面,它与B细胞表面的CD40的结合为B细胞提供第二活化信号。
CD40L-CD40的配接不但与B细胞的活化有关,而且与抗体的转类有关。
CD40传送的信号诱导B细胞中B7分子表达的上调,反过来又为Th细胞提供了第二信号。
7.丝裂原受体
丝裂原(mitogens)是一类能够非特异性地激活T或B淋巴细胞的因子。
因为它们能同时激活许多淋巴细胞克隆,所以丝裂原被称为多克隆激活剂(polycolonalactivators)。
常见的能激活T细胞的凝集素包括植物血凝素(PHA)和刀豆蛋白A(ConA)。
8.粘附分子
T细胞表面表达许多粘附分子,这些粘附分子与其它细胞表面相应的粘附分子的相互作用参与T细胞的活化、移动和效应等功能。
T细胞表达的选择素与细胞的移动和归巢有关。
CD2(LFA-2),LFA-1分别与APC或靶细胞表面ICAM-1(CD54),LFA-3(CD58)等结合,延长并加强T细胞与其他细胞之间的接触,促进TCR对MHC分子-肽的识别,同时为T细胞活化提供第二信号。
(三)T细胞亚群
1.abT细胞和gdT细胞
这两类T细胞分别表达abTCR和gdTCR。
abT细胞占胸腺和外周血中T细胞的大多数。
但是在粘膜上皮中,大多数是gdT细胞。
这些粘膜gdT细胞的特征是表达CD8,而外周血中的gdT细胞大多数为双阴性细胞,不表达CD8。
粘膜中CD8+gdT细胞的多样性不如abT细胞丰富,它们的TCR主要针对某些细菌和病毒抗原,在粘膜表面发挥保护作用。
此外,某些gdT细胞能直接识别抗原而不需要APC递呈。
2.CD4+和CD8+T细胞
CD4+T细胞在功能上主要是辅助细胞(Th细胞),而CD8+T细胞在功能上主要是细胞毒性细胞(CTL)。
CD4分子和CD8分子限定了T细胞所能识别的MHC分子的类型:
CD4+T细胞识别MHCII类-肽复合物,CD8+T细胞识别MHCI类-肽复合物。
3.Th1和Th2细胞亚群
根据所分泌的细胞因子格局的不同,CD4+T细胞可进一步分为Th1和Th2两个亚群。
Th1和Th2产生的细胞因子种类不同,这是鉴别这两种细胞亚群的主要依据,也是两个亚群功能差别的主要基础。
Th1细胞主要产生IL-2、IFN-g和TNF-b,而Th2主要产生IL-4和IL-5。
Th1参加迟发型超敏反应,辅助CD8+T细胞活化,促进Mf活化和促进IgG2a的产生。
所以Th1细胞主要参与细胞介导的炎症反应。
Th2细胞主要辅助B细胞对胸腺依赖性抗原的应答,刺激肥大细胞和嗜酸性粒细胞生长和分化。
因为IL-4促进IgE的转类,所以Th2细胞促进I型变态反应的发生。
Th1和Th2在功能上互相拮抗。
Th1分泌的细胞因子抑制Th2的功能,同样地,Th2分泌的细胞因子抑制Th1的功能。
4.初始T细胞和记忆T细胞
初始T细胞指离开胸腺后,从未接触过抗原的成熟T细胞。
初始T细胞是静止的、处于G0期的细胞。
初始T细胞集中在外周淋巴器官和组织中的T细胞区内。
初始T细胞的标志是CD45RA。
记忆T细胞是指已经接触过抗原(已致敏)但是没有经历终末分化成为效应细胞的T细胞,其标志是CD45RO。
记忆T细胞是长寿细胞,存在于血液和外周淋巴组织和器官中。
记忆细胞在再次遇到相同抗原时,产生迅速而又强烈的反应。
刺激产生记忆细胞(包括T和B记忆细胞)是疫苗接种的重要基础之一。
二、B淋巴细胞
(一)B淋巴细胞发育成熟
B细胞成熟过程分骨髓内和骨髓外两个阶段。
本处叙述的B细胞均指B2细胞,不包括B1细胞。
骨髓内分化包括从淋巴干细胞发育成为成熟B细胞,这一发育阶段不依赖抗原,是BCR基因重排和对重排基因产物选择的过程,此外还有其他表面分子表达的变化。
B细胞骨髓外发育受抗原驱动,在外周淋巴器官滤泡的生发中心内进行,包括亲和力成熟、Ig转类以及浆细胞和记忆B细胞形成三个事件。
B细胞骨髓外的发育实际上是B细胞在对胸腺依赖性抗原的免疫应答过程中发生的Ig基因的体细胞高频突变、对高亲和力受体的选择和重链基因重组的过程。
也可以说是在非己抗原驱动下,抗原受体进一步发育和被选择的过程,使得所产生的抗体更适合于与抗原结合。
1.抗体亲和力成熟
亲和力成熟是指抗体平均亲和力的提高,通常在再次应答中发生产生。
B细胞对抗原产生应答后,其BCR基因发生高频突变,产生高亲和力BCR的B细胞被选择,产生低亲和力BCR的B细胞被淘汰,所以抗体的亲和力提高。
2.抗体类别转换
体细胞高频突变后发生Ig重链同种型转化重组,g、a和e重链取代m链,分别产生IgG、IgA或IgE类抗体。
转类过程中抗体分子的Fab段保持不变,转类后的抗体的特异性因此也保持不变。
由于抗体的生物学效应是由Fc段决定的,转类可以改变抗体的生物学效应。
转类需要Th2细胞的辅助和细胞因子的参与。
3.浆细胞和记忆细胞的分化
表达高亲和力受体的中心母细胞分化成为浆细胞和记忆细胞。
(二)B细胞表面分子及功能
这里叙述的是成熟B细胞表达的各种膜分子,它们在B细胞的功能中起不同的作用。
1.BCR和BCR复合体
B细胞的抗原受体即是成熟B细胞膜表面的IgM和IgD。
IgM和IgD与另外两条链Iga和Igb一起组成BCR复合体,
Iga-Igb是BCR的装配和在细胞膜上的表达所必需的,并且转导BCR识别抗原后产生的第一信号。
2.BCR协同受体
与TCR一样,BCR也有协同受体。
BCR的协同受体包括CD19,CD21(CR2)和CD81(TAPA-1)3种膜分子,它们在B细胞膜上与BCR直接接触(图9-8)。
BCR协同受体能够促进通过BCR-Iga-Igb产生的活化信号。
BCR协同受体的参与可使BCR复合体介导的信号提高1000至10000倍。
3.CD40
CD40是B细胞膜上表达的一种糖蛋白,它与CD40L结合后向细胞发出第二信号,导致B细胞激活、增殖并分化。
CD40与CD40L还在抗体转类中起重要作用。
。
CD45已在T淋巴细胞中叙述,它也在B细胞表面表达。
5.B7
B7包括B7-1(CD80)和B7-2(CD86)。
B7-1和B7-2以同源二聚体形式表达在B细胞表面,是APC表面最重要的协同刺激分子。
静止B细胞不表达B7,B细胞在摄入抗原后诱导性表达B7。
Th细胞表面CD40L与B细胞表面CD40相互作用可使B7的表达上调。
B7是T细胞表面的CD28和CTLA-4的配体(图9-3)。
B7与CD28的相互作用为T细胞提供第二活化信号,延长和增强IL-2和其他细胞因子的产生,阻止耐受的发生,并促进CD40L的表达。
B7与CTLA-4的结合抑制IL-2受体a链的表达,抑制IL-2的分泌和IL-2mRNA的积累,抑制细胞分裂,其结果是抑制T细胞继续活化。
6.MHC
B细胞表达MHCI类和II类分子,MHCII类分子的作用是向T细胞递呈外源性抗原。
金葡菌A蛋白(SPA)能使B细胞分裂。
小鼠B细胞表面有革兰阴性菌细胞壁的脂多糖(LPS)受体。
脂多糖中起丝裂原作用的是其脂类成分。
8.细胞因子受体
活化B细胞表达IL-4、-5、-6等细胞因子的受体,在Th2细胞分泌的相应的细胞因子的作用下,增殖和分化。
(三)B细胞亚群
根据发育早晚、存在部位、表面标志和功能的不同,可以把B细胞分成B1和B2两个亚群。
B1细胞组成性表达CD5,所以又称为CD5+B细胞,B2细胞不表达CD5,故称为CD5-B细胞,但是活化的B2细胞也表达CD5。
1.B1细胞
B1细胞主要存在于体腔表面,例如肠道粘膜的固有层。
其表面表达IgM而不表达IgD。
主要产生IgM类抗体。
B1细胞的多样性贫乏,只产生胚系抗体基因编码的抗体。
这些抗体的特异性差,交叉反应强(polyreactive),而且亲和力低。
B1细胞主要识别微生物的多糖和脂类抗原,这种抗体应答不依赖T细胞的辅助。
B1细胞主要有以下三方面功能:
第一,在体腔表面发挥抗微生物作用。
第二,通过产生自身抗体发挥免疫自稳作用。
B1细胞产生的自身抗体清除衰老细胞,维持免疫耐受。
第三,独特型网络的调节作用。
2.B2细胞
如果不加说明,通常所指的B细胞就是B2细胞。
B2细胞介导抗胸腺依赖性抗原体液免疫,其特征已在前面叙述。
0.免疫球蛋白的功能:
1.抗体Ig分子可看作是一个四方面的“二合一”分子:
结构上:
由重链和轻链组成、由Fab和Fc组成;
功能上:
Fab段/V区表现抗原识别功能,Fc段/C区表现效应功能;
特性上:
既是抗体—与抗原结合、又是一个抗原—被抗抗体结合;
效应上:
既可对机体有利,也可对机体有害。
2.IgV区的功能-----特异性结合抗原:
在体内,可中和毒素、阻断病原入侵、清除病原微生物或导致免疫病理损伤;
B细胞膜表面的IgM和IgD构成B细胞的抗原识别受体,能特异性识别抗原分子;
在体外可发生各种抗原抗体结合反应,有利于抗原或抗体的检测和功能的判断;
0.IgG:
1.再次免疫应答产生的主要抗体,亲和力高,分布广;
2.是机体抗感染的“主力军”;
3通过胎盘(IgG1、IgG2、IgG3,新生儿抗感染);
4.激活补体(裂解细胞);
5.调理作用(促进吞噬);
6.介导ADCC(细胞毒作用)
3.参与T细胞识别、粘附及活化的CD分子的种类、结构特点、识别配体及其功能有:
种类
结构特点
识别配体
功能
CD3
五聚体,与TCR组成TCR/CD3复合物
稳定TCR结构、传递活化信号
CD4
单体分子
MHCⅡ类分子
增强TCR与APC或靶细胞的亲和性,并参与信号传导。
CD8
异源二聚体
MHCⅠ类分子
CD2
CD58(LFA-3)
增强T细胞与APC或靶细胞的粘附及CD2分子所介导的信号传导
CD58
促进T细胞识别抗原,参与T细胞信号传导
CD28
同源二聚体
B7
提供T细胞活化的辅助信号
CD152
对T细胞活化有负调节作用
CD40L
三聚体
CD40
是B细胞进行免疫应答和淋巴结生发中心形成的重要条件
4.参与B细胞识别、粘附及活化的CD分子的种类、结构特点、识别配体及其功能有:
CD79
与mIg组成BCR复合物,介导B细胞信号传导
CD19
促进B细胞激活
CD21
C3片段EB病毒
增强B细胞对抗原的应答,参与免疫记忆
CD80/CD86
CD40
0.白细胞分化抗原的生物学作用有:
参与细胞生长、分化、正常组织结构的维持
参与免疫应答过程中免疫细胞的相互识别,免疫细胞抗原识别、活化、增值和分化,以及免疫功能的发挥
造血细胞的分化和造血过程的调控
参与炎症的发生、血栓形成和组织修复
肿瘤的恶化和转移。
0.试述免疫应答的基本过程?
免疫应答的过程可人为地划分为三个阶段:
即识别阶段、活化与分化阶段、效应阶段。
识别阶段:
抗原被APC所摄取、加工、处理;
T细胞/B细胞通过TCR/BCR特异性识别抗原肽。
提呈和识别的过程。
活化与分化阶段:
T、B细胞特异性识别抗原,产生其活化的第一信号;
T/B细胞与APC表面的多种粘附分子间相互作用,提供T细胞活化的第二信号(即协同刺激信号)。
另外,激活的APC和T细胞产生多种细胞因子,它们以自分泌和旁分泌作用,参与淋巴细胞的增殖与分化,最终形成T效应细胞或奖细胞,并分泌免疫效应分子如各种细胞因子和抗体。
效应阶段:
效应细胞和效应分子共同发挥作用,清除非己抗原物质或诱导机体产生免疫耐受,从而维持机体正常生理状态,或引起免疫相关疾病。
0.简述B细胞接受抗原刺激后分化成熟的过程及转归:
B细胞直接识别天然抗原表位,无需APC对抗原的处理和呈递。
接受抗原刺激的特异性B细胞,进入外周淋巴组织后与相应的抗原特异性Th细胞相遇,被阻留在T细胞区,接受Th2细胞的辅助并活化,活化后,一部分B细胞迁移至髓质,分化为浆细胞,产生IgM类抗体(IgM类为主),两周后凋亡;
另一部分B细胞迁移至附近的B细胞区(初级淋巴滤泡),继续增殖并形成生发中心(次级淋巴滤泡)。
在生发中心,一部分B细胞分化为浆细胞,迁移至骨髓,产生高亲和力抗体(IgG类为主),还有部分分化为记忆B细胞,参与再次免疫应答。
B细胞在生发中心分化成熟过程中经历三个事件:
即体细胞高频突变和Ig亲和力成熟、抗原受体编辑及抗体类别转化。
0.细胞黏附分子的功能:
1炎症过程中白细胞与血管内皮细胞的黏附2黏附分子参与淋巴细胞的归巢3黏附分子参与免疫细胞的识别与活化4黏附分子参与细胞的发育、分化、附着及迁移5黏附分子与肿瘤细胞的


- 配套讲稿:
如PPT文件的首页显示word图标,表示该PPT已包含配套word讲稿。双击word图标可打开word文档。
- 特殊限制:
部分文档作品中含有的国旗、国徽等图片,仅作为作品整体效果示例展示,禁止商用。设计者仅对作品中独创性部分享有著作权。
- 关 键 词:
- 免疫学 笔记 复习 知识点 总结
 冰豆网所有资源均是用户自行上传分享,仅供网友学习交流,未经上传用户书面授权,请勿作他用。
冰豆网所有资源均是用户自行上传分享,仅供网友学习交流,未经上传用户书面授权,请勿作他用。
 对中国城市家庭的教育投资行为的理论和实证研究.docx
对中国城市家庭的教育投资行为的理论和实证研究.docx
-
二年级下册数学练习题大全.docx
-
二十年后回故乡的优秀作文.docx
-
软基换填施工方案.docx
-
《黑白装饰画》教案.docx
-
课堂教学改革实施方案5篇.docx
-
返璞归真简约致美解读《给予树》教学设计语文.docx
-
离职证明范本精选多篇.docx
-
《天局》全文.docx
-
我害怕作文集合15篇.docx
-
伏魔战记39详细攻略.docx
-
幼儿园学期计划.docx
-
雅思分类打印版Word格式文档下载.docx
-
年产1万吨竹子纤维加工项目可行性研究报告文档格式.docx
-
电商产业化项目投资经营商业计划书Word文件下载.docx
-
医学多媒体课件的设计与制作Word文档格式.docx
-
中学生中秋节想象作文Word格式.docx
-
等保20之漏洞扫描系统技术方案建议书Word文档格式.docx
-
培训学校个人工作计划模板5篇Word格式.docx
-
北京各区二模试题分类汇编文言文阅读Word文档下载推荐.docx
-
不同职业病危害因素的防护常识Word格式文档下载.docx
-
一年级上册同音形近字练习汇总Word文档格式.docx
-
班级家长会上班主任教师讲话稿Word下载.docx
-
科斯塔环载波恢复Word文件下载.docx
-
浙教义务版六年级语文下册教案 花潮Word文件下载.docx
-
集成电路设计与集成系统专业Word格式文档下载.docx
-
开工第一课专题讲座观后感文档格式.docx
-
东城区学年第一学期高三期末化学试题及答案Word格式文档下载.docx
-
苏教版六年级语文下册第七单元测试题Word格式文档下载.docx
-
学长征精神做红色传人活动方案文档格式.docx
-
读书笔记150字30篇文档格式.docx
-
中级经济法考前必背法条精华版备考资料Word格式.docx
-
国家公务员考试《行政职业能力测验》真题卷B类参考答案与解析.docx
-
工程监理年终个人工作总结5篇.docx
-
公共交通数据仓库平台及数据挖掘项目说明书IBM.docx
-
过秦论读后感.docx
-
公众应急手册.docx
-
高考英语真题分类汇编专题03非谓语动词和交际用语解析版.docx
-
工业产品CAD技能三级试题及其评分标准题精.docx
-
寒假作业.docx
-
固体废弃物加工免烧砖项目建设可行性研究报告固体废物综合利用项目.docx
-
高三理综上学期第一次月考试题1.docx
-
好词大全2字和4字.docx
-
公共部门危机管理中的预警机制建设.docx
-
关于某中心各装置现场整治实施细则之消漏讲解.docx
-
高一化学必修一期中试题整理.docx
-
公务员年度考核个人总结5篇.docx
-
管道技术标详解资料.docx
-
高一政治我国政府受人民的监督测试题.docx
-
古仙界民俗民情旅游开发建设项目可研技术方案.docx
-
广东工业大学计算机网络实验报告.docx
