 TGF信号通路.docx
TGF信号通路.docx
- 文档编号:18184703
- 上传时间:2023-04-24
- 格式:DOCX
- 页数:9
- 大小:1.21MB
TGF信号通路.docx
《TGF信号通路.docx》由会员分享,可在线阅读,更多相关《TGF信号通路.docx(9页珍藏版)》请在冰豆网上搜索。
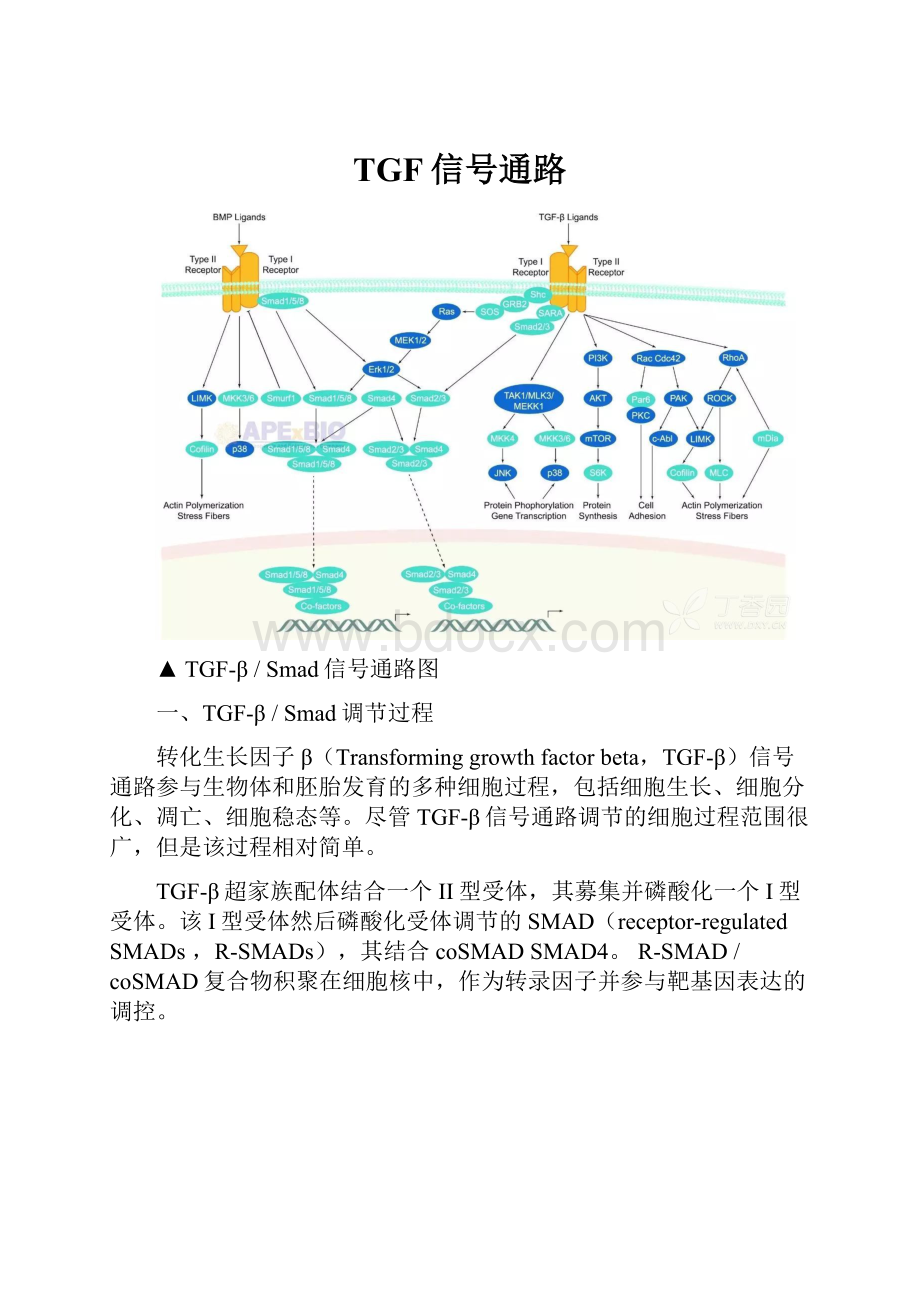
TGF信号通路
▲TGF-β/Smad信号通路图
一、TGF-β/Smad调节过程
转化生长因子β(Transforminggrowthfactorbeta,TGF-β)信号通路参与生物体和胚胎发育的多种细胞过程,包括细胞生长、细胞分化、凋亡、细胞稳态等。
尽管TGF-β信号通路调节的细胞过程范围很广,但是该过程相对简单。
TGF-β超家族配体结合一个II型受体,其募集并磷酸化一个I型受体。
该I型受体然后磷酸化受体调节的SMAD(receptor-regulatedSMADs,R-SMADs),其结合coSMADSMAD4。
R-SMAD/coSMAD复合物积聚在细胞核中,作为转录因子并参与靶基因表达的调控。
TGF-β配体结合受体
TGF-β超家族配体包括:
骨形态发生蛋白(Bonemorphogeneticproteins,BMPs),生长和分化因子(Growthanddifferentiationfactors,GDFs),抗缪勒氏管激素(Anti-müllerianhormone,AMH),激活素(Activin),Nodal和TGF-β。
配体分为两个分支:
(1)TGF-β/激活素分支和
(2)BMP/GDF分支。
TGF-β/激活素分支包括TGF-β,激活素,Nodal。
BMP/GDF分支包括BMP,GDF和AMH配体。
信号传导从TGF-β超家族配体与TGF-βII型受体的结合开始。
II型受体是丝氨酸/苏氨酸受体激酶,其催化I型受体的磷酸化。
每种类型的配体结合特异性II型受体,在哺乳动物中有七种已知的I型受体和五种II型受体。
II型受体募集I型受体和磷酸化
TGF-β配体结合II型受体二聚体,其募集I型受体二聚体,与配体形成杂四聚体复合物。
II型受体磷酸化I型受体的丝氨酸残基。
I型受体磷酸化R-SMAD
有5个受体调节的SMAD:
SMAD1,SMAD2,SMAD3,SMAD5和SMAD9(有时称为SMAD8)。
TGF-β,激活素,Nodal和一些GDFs由SMAD2和SMAD3介导,而BMP,AMH和少量GDFs由SMAD1,SMAD5和SMAD9介导。
R-SMAD与I型受体的结合由含锌双指FYVE结构域的蛋白质介导。
介导TGF-β途径的两种这样的蛋白质包括SARA(Smad受体激活锚定蛋白)和HGS(肝细胞生长因子调节酪氨酸激酶底物)。
CoSMAD结合
磷酸化的RSMAD对coSMAD(例如SMAD4)具有高亲和力,并与其形成复合物。
磷酸盐基团不作为coSMAD的对接位点,而是磷酸化开启了允许相互作用的氨基酸延伸。
R-SMAD-coSMAD复合物进入细胞核
磷酸化的RSMAD/coSMAD复合物进入核,结合转录启动子/辅因子并引起DNA的转录。
骨形态发生蛋白导致涉及成骨,神经发生和腹侧中胚层特化的mRNA转录。
TGF-β导致参与凋亡,细胞外基质新生和免疫抑制的mRNA转录。
它也参与G1细胞周期的停滞。
激活素引起参与性腺生长,胚胎分化和胎盘形成的mRNA转录。
Nodal引起左轴和右轴特化,中胚层和内胚层诱导中涉及的mRNA转录。
二、相关蛋白或基因
1.SMAD
SMAD是细胞内蛋白,其将细胞外信号从转化生长因子β配体转移到细胞核,激活下游基因转录。
SMADs是I型受体唯一的底物。
SMADs分为三个亚族:
受体调节SMADs(R-SMADs:
SMAD1,SMAD2,SMAD3,SMAD5,SMAD8),被I型受体磷酸化;常用介体SMADs(Co-SMADs:
SMAD4),其与活化的R-SMADs低聚;和抑制性SMADs(I-SMADs:
SMAD6和SMAD7),其由TGF-β家族成员诱导。
SMAD6和SMAD7作为抑制性SMAD(I-SMADS),它们在调节TGF-β信号中发挥关键作用,并参与负反馈。
SMAD7与R-SMADs竞争,并阻止I型受体的磷酸化。
SMAD6结合SMAD4,阻止其它R-SMAD与coSMAD的结合。
SMADs具有两个保守结构域,N末端Mad同源1(MH1)和C末端Mad同源2(MH2)结构域。
MH1结构域通过与DNA结合并与核蛋白质相互作用来调节核进入和转录,MH2结构域调节SMAD寡聚化。
在由I型受体磷酸化R-SMADs之后,认为发生SMAD低聚。
未磷酸化的SMAD蛋白主要作为单体存在,并且在磷酸化时,R-SMADs形成同源寡聚体,其快速转化成含有Co-SMAD(SMAD4)的异寡聚体。
早期实验证实低聚SMADs是三聚体,后来有人提出SMAD2-SMAD4复合物的二聚体构型。
2.Activin&inhibin
激活素和抑制素。
是两个密切相关的蛋白质复合物,生物学效应几乎相反。
激活素能增强FSH的生物合成和分泌,并参与月经周期的调节。
相反,抑制素下调FSH合成并抑制FSH分泌。
激活素是由两个相同或非常相似的β亚基构成的二聚体。
抑制素也是二聚体,其中第一组分是与激活素中的β亚基相似或相同的β亚基。
然而,与激活素相反,抑制素二聚体的第二个成分是一个相关性远一点的α亚基。
均属于TGF-β蛋白超家族。
3.GDFs
Growthdifferentiationfactors,生长分化因子。
GDF1主要表达在神经系统中,在胚胎发育中起作用。
GDF2(也称为BMP9)诱导和维持胚胎基底前脑胆碱能神经元(BFCN)对神经递质乙酰胆碱的反应,并通过增加铁调素蛋白质的水平调节铁代谢。
GDF3也称为“Vg相关基因2”(Vgr-2)。
它具有双重功能,抑制和诱导胚胎发育的早期阶段。
GDF5在发育的中枢神经系统中表达,在关节和骨骼的发育中起作用,并增加响应多巴胺的神经元的存活。
GDF6与骨形态发生蛋白相互作用,调控眼睛发育。
GDF8现在被称为肌肉生长抑制素并控制肌肉组织的生长。
GDF9,像GDF3,相对于TGF-β超家族的其它成员缺乏一个半胱氨酸。
其基因表达限于卵巢,并且在排卵中起作用。
GDF10与BMP3密切相关,在头部形成中起着重要作用,它也被称为BMP-3b。
GDF11调节嗅觉上皮中发生的嗅觉受体神经元的数量和视网膜中发育的视网膜神经节细胞的数量。
GDF15(也称为TGF-PL,MIC-1,PDF,PLAB和PTGFB),在组织损伤和某些疾病过程中具有调节炎症和凋亡途径的作用。
4.BMP
Bonemorphogeneticproteins,骨形态发生蛋白。
最初发现了七种这样的蛋白质。
其中6种(BMP2〜BMP7)属于TGF-β超家族。
BMP1是金属蛋白酶。
此后,又发现了十三个BMPs,总数达二十个。
5.Nodal
Nodal是一种分泌蛋白,属于转化生长因子(TGF-β)超家族。
它受到抑制因子LeftyA,LeftyB,Cerberus和Tomoregulin-1的严格调控,可以干扰Nodal受体的结合。
6.Lefty
Lefty(left-rightdeterminationfactors),是新近发现的TGF-β超家族成员之一。
这些蛋白质是分泌的,并在发育过程中器官系统的左右不对称性测定中发挥作用。
编码这些蛋白质的基因的突变与左右轴畸形相关,特别是在心脏和肺中。
是Nodal信号途径的拮抗剂。
7.TGF-β
是属于转化生长因子超家族的多功能细胞因子,包括四种不同的同种型(TGF-β1~4)。


- 配套讲稿:
如PPT文件的首页显示word图标,表示该PPT已包含配套word讲稿。双击word图标可打开word文档。
- 特殊限制:
部分文档作品中含有的国旗、国徽等图片,仅作为作品整体效果示例展示,禁止商用。设计者仅对作品中独创性部分享有著作权。
- 关 键 词:
- TGF 信号 通路
 冰豆网所有资源均是用户自行上传分享,仅供网友学习交流,未经上传用户书面授权,请勿作他用。
冰豆网所有资源均是用户自行上传分享,仅供网友学习交流,未经上传用户书面授权,请勿作他用。
 《地质测量图绘制细则》.docx
《地质测量图绘制细则》.docx
-
《大学生健康教育教程》考试题库500题DR含答案.docx
-
《黄鹤楼送孟浩然之广陵》字词注释.docx
-
《葱爆羊肉》概要.docx
-
《面向对象程序设计》C++综合练习学生版.docx
-
《化学反应工程》课程综合复习资料.docx
-
《社会心理》网上形考参考答案.docx
-
《轮扣式支架》模板施工方案.docx
-
《团委新成员大会主持稿1》.docx
-
《三峡学典》思品七上.docx
-
《有效备课上课听课评课》读书笔记.docx
-
《土地的誓言》导学案.docx
-
1汽车维修专业人才需求调研报告.docx
-
《应用文写作》教学设计.docx
-
1级三相费控智能电能表技术规范.docx
-
17#楼机房层模板施工方案.docx
-
《稻草人》精彩读书笔记精选多篇doc.docx
-
《机械制造工艺与装备》复习资料.docx
-
《木兰从军》教案.docx
-
《神奇的毛细现象》.docx
-
《围城》读书心得精选范文1000字5篇.docx
-
《幼儿园是我家》大班教案.docx
-
1资产负债表变动情况的分析评价.docx
-
5s活动总结报告5s总结报告2篇.docx
-
《当代学生》初中文言文阅读训练篇.docx
-
《机械制造工艺学》电子教案.docx
-
《民事案件案由规定》物权纠纷案由的理解与适用.docx
-
《社会学研究方法学习心得5篇范文》.docx
-
《外商投资企业联合年检报告书》填写要求和指标说明.docx
-
《幼儿园课程论》形成性考核册答案27497.docx
-
1在直接影响物价水平的诸多因素当中.docx
-
10套普通话练习材料.docx
-
JAVA国际化Word下载.docx
-
北京市第五届职业技能竞赛安全应急领域应急救援员水域搜索与救援比赛竞赛大纲模板Word文件下载.docx
-
Linux网络配置Samba文档格式.docx
-
北师大版小学语文五年级下册教学计划1Word文档格式.docx
-
变压器状态检修问题探讨Word格式.docx
-
财务分析考题问答版Word下载.docx
-
SQL Server 安装和配置图解Word下载.docx
-
测量Word文档下载推荐.docx
-
初中文言文实词虚词积累文档格式.docx
-
弟子规全文带拼音完整版Word格式文档下载.docx
-
第七章 地理信息系统工程与应用Word格式文档下载.docx
-
典型案例学习材料Word下载.docx
-
原创菏泽高考模拟最新试题Word文件下载.docx
-
助学贷款演讲稿文档格式.docx
-
浙江省建筑节能验收规范含资料表格文档格式.docx
-
职业健康安全系统管理系统方案设计00916文档格式.docx
-
资料员最新考试试题和答案解析Word格式文档下载.docx
-
中国古代文化常识Word下载.docx
-
自考人力资源管理一资料Word文档格式.docx
