 大一普通生物报告文档格式.docx
大一普通生物报告文档格式.docx
- 文档编号:17135676
- 上传时间:2022-11-28
- 格式:DOCX
- 页数:17
- 大小:1.61MB
大一普通生物报告文档格式.docx
《大一普通生物报告文档格式.docx》由会员分享,可在线阅读,更多相关《大一普通生物报告文档格式.docx(17页珍藏版)》请在冰豆网上搜索。
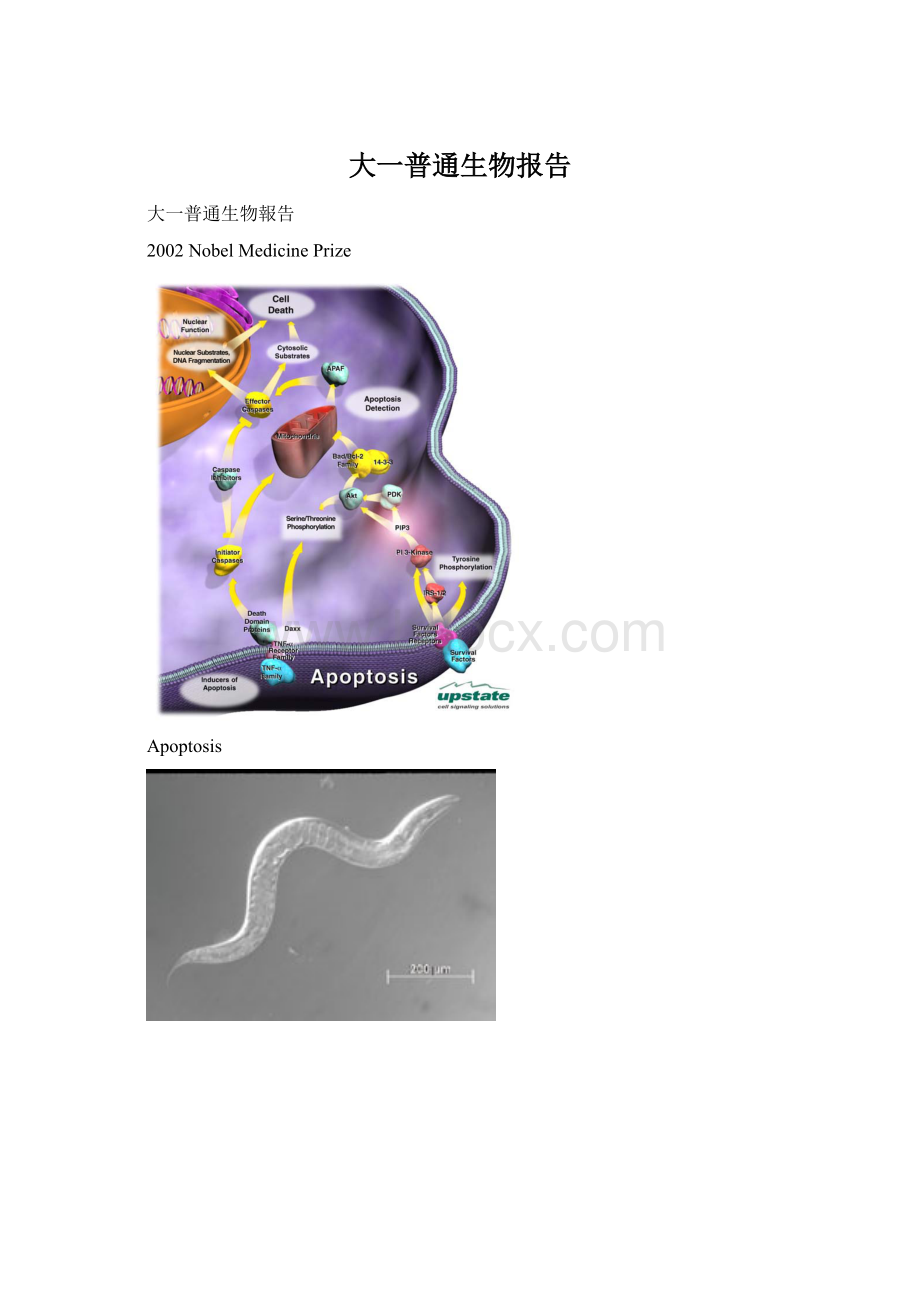
首先來看發育,從神經發育的過程來架構整個神經細胞的生存與死亡。
在神經發育的過程裡,為了要確定未來在神經系統的組成是足夠的,細胞在剛開始製造的細胞數是後來成熟神經系統的2-3倍數量。
在神經系統成熟的過程中,經由細胞死亡,修飾成完整而有功能的神經系統。
而有些動物在發育的過程中是必須經過apoptosis,例如人類的手指和青蛙的尾巴。
在人體的新陳代謝中,人體的細胞也在不斷更新。
人體內細胞的誕生和死亡處於一個動態平衡階段,一個成年人體內每天都有上萬億細鮑誕生,同時又有上萬億細胞「計畫性死亡」兩者處於一種動態平衡中,體器官維持合適的細胞數量得以正常運作的,正是「計畫性細胞死亡」機制。
Apoptosis也與維持環境恆定有關係,例如t細胞在胸線中發育不完全的話,就會自動啟動apoptosis,避免t細胞殺掉不該殺的細胞。
Apoptosis如果發生異常狀況的話,會導致疾病,例如癌症,即是當細胞發生突變時,apoptosis未正常發揮作用,使細胞凋亡,就會使突變細胞在進一步的突變,產生原發性腫瘤或轉移性腫瘤。
而愛滋病,起因為apoptosis殺死不該殺死的細胞。
人類的手指經由apoptosis而發育完成
Cellcycle圖
當cellcycle在進行細胞複製時,出現問題而被checkpoint檢查到,細胞中的p53就會被啟動,然後中止細胞分裂,試著去修補受損的鹼基對,如果修補好了,則p53就會失去作用,使細胞繼續進行細胞複製的行程,若修補不好,則p53不斷累積,達到一個定量後,就會啟動apoptosis。
凋亡
壞死
機制
基因調控的程序化細胞死亡,主動進行
意外事故性細胞死亡,被動進行
死亡範圍
多為散在的單個細胞
多為聚集的大片細胞
形態特徵
細胞固縮,核染色質邊聚或中聚,內質網擴張,凋亡細胞皺縮,與周圍細胞失去結合並脫落,膜可發泡成芽,形成凋亡小體
細胞腫脹,核染色質絮狀或邊集,細胞膜及各細胞器膜溶解破壞,溶酶體酶釋放,細胞自溶
細胞內部生理反應
耗能的主動過程,有新蛋白合成,DNA早期規律降解為180-200bp片段,凝膠電泳呈特徵性梯帶狀
不耗能的被動過程,無新蛋白合成,DNA降解不規律,片段大小不一,凝膠電泳不呈梯帶狀
凋亡
壞死
周圍反應
不引起周圍組織炎症反應和修復再生,但凋亡小體可被鄰近細胞吞噬
引起周圍組織炎症反應
細胞核
被細胞凋亡所產生的蛋白質將核膜切割形成核小體,切割處位於核孔,此外核內的核纖維體也被水解
1.核濃縮
2.核碎裂
3.核溶解
細胞骨架
被細胞凋亡所產生的蛋白質給水解
此圖為apoptosis的dna降解後拿去電泳所產生的ladder
左邊為necrosis右邊為apoptosis
左邊的圖為凋亡小體右圖為apoptosis的細胞,apoptosis細胞通常為單一的細胞。
程式性細胞死亡-細胞自噬
細胞自噬是一種經由溶體來降解細胞內的胞器,他在細胞生長發展跟新陳代謝都有影響,他的主要機制為在一個缺乏營養的細胞中,將營養從不需要的區域分配到需要的地方。
有人提出因為細胞自噬因而導致細胞死亡的過程為一種程式性的細胞死亡,但並沒有證據,但是有觀察到程式性細胞死亡的細胞出現細胞自噬的現象,所以有autophagiccelldeath這個詞的出現。
根據研究顯示昆蟲在蛻變時出現了一種計劃性細胞死亡,但是與其他種程式性細胞死亡不同,被認為是autophagiccelldeath。
現在還不確定自噬現象是導致細胞死亡的主因還是只是細胞死亡過程中的一部分。
自噬過程中,細胞需要一個將有待分解的物質包圍並隔離起來,叫做自噬體,由雙層脂質膜構成,然後再與lysosome結合溶掉不需要的胞器(老化或病死)。
得獎人介紹
今年諾貝爾生理及醫學獎頒給加州柏克萊大學布睿能(SydneyBrenner)博士、麻省理工學院霍維茲(RobertHorvitz)博士及英國劍橋大學桑格中心蘇斯坦(JohnE.Sulston)博士,肯定三位在遺傳基因調控於器官發育及細胞死亡上的卓越貢獻。
在1961年Brenner博士與賈柯與麥瑟森一同發現了mRNA,同年Brenner與克立克(也就是發現雙股螺旋的人之一),一起發現了codentriplet-三個核甘酸可以對應一個胺基酸。
1963年Brenner提出了線蟲做為動物模式(animalmodel)-有簡單結構和易於研究觀察的動物,為何要用線蟲做為animalmodel?
是因為線蟲為非寄生性的生物,構造簡單,成蟲的細胞只有959個,相對於常用來當作實驗材料的果蠅有十萬個細胞,果蠅筆線蟲簡單的多,而且很容易培養,生命週期短,可像微生物般操作,也可冷凍保存,且全身透明,易於解剖與觀察。
1967年發現EMS(ethylmethanesulphonate)又發現蟲產生單點突變,利用分析這些突變株,就可遭受破壞的基因在胚胎發育過程中所擔任的功能。
1974年Brenner發表第一篇有關線蟲遺傳學操作原理的論文,裡頭詳述他分離出約300種的細蟲突變株,並分析這些突變株的表現型。
1983年Sulston發表線蟲的細胞淵源(celllineage)並找到了參與細胞凋亡的基因-nuc-1,細胞淵源就是細胞分裂和細胞親緣的關係,也就是討論野生種內的每個細胞何時分裂,分裂成幾個細胞,分裂後的細胞命運,當我們對線蟲的每個細胞都有了了解,我們才能找出有變異的突變種,也可對細胞分裂的機制加以研究。
Sulston所作的細胞淵源實驗,是說明了一個受精卵可以發育成1090個,而成蟲只有959個細胞,另外
131個體的細胞會在特定發育時期,進行計劃性細胞死亡。
Nuc-1表現為DNase,和死細胞內的DNA分解有關。
1986年Horvitz找到了與計劃性細胞死亡有直接關係的基因-ced-4、ced-3,若ced-3、ced-4發生了突變,幾乎沒有一個線蟲細胞可以進行計劃性細胞死亡,之後Horvitz又找到另一基因ced-9,這個基因藉由抑制ced-3、ced-4的作用,扮演保護細胞的角色,避免細胞進行細胞死亡。
1993年Horvitz發現線蟲的ced-3所表現的蛋白質是cysteineprotease,此蛋白相似於哺乳類的caspase。
死亡訊息的傳遞路徑
細胞凋亡的「執行者」,是為Cascade3的protein,它可以用來活化DNase,進而切割DNA,也可直接分解結構性蛋白,如lamin、actin;
但是當死亡訊息到來的時候,Cascade3要怎麼接收到並且加以執行,這是以下要討論的題目。
當死亡訊息到來的時候,其實有兩類主要的路徑,一是透過細胞膜受體的訊息傳遞路徑,而另一是透過粒線體來發動。
線蟲(C.elegans)
很多人一聽到線蟲都會以為是對人體有害的寄生蟲,其實不然,線蟲對人體無害,生長於土壤之中,以生物碎屑和一些微生物為食物。
線蟲全長大約一毫米,以人體肉眼勉強看的到,對於做為動物模式(animalmodel)有著非常多優點,由左圖可知線蟲體透明,所以非常方便觀察,而且在實驗室培養相當容易,在室溫攝氏25度即可培養,而且線蟲的生活史只有3天半,生長相當快,線蟲又分為雌雄同體(右圖上)及雄性(右圖下),而在實驗室正常狀態培養出來的絕大多數屬於雌雄同體,雌雄同體的線蟲行體內授精,所以後代與親代基因一模一樣,可以維持基因純品性,一隻雌雄同體的線蟲可以產生200到300隻後代,正常培養一個禮拜下來,大約會產生一萬隻線蟲,因為其體透明,細胞分裂又有一定速度,而且每個細胞相對位置都固定,可以直接用顯微鏡追蹤觀察,今天觀察幾個,明天可以繼續觀察下去不用重複今天觀察的,有人把這種優點比喻成看小說,今天看到第15頁,明天可以直接從第16頁看起,不需要從頭再看,以上列舉的優點都是科學家選線蟲為動物模式(animalmodel)的原因。
線蟲的細胞凋亡
由圖知一開始健康的細胞要執行細胞凋亡,而形成死細胞,死細胞之後又被吞噬細胞所吞噬,最後DNA被分解掉,裡面直接跟細胞凋亡有關的基因為ced-9、ced-3、ced-4,其中ced-3是直接造成細胞凋亡的基因,ced-4目的是去活化ced-3進一步執行細胞凋亡,所以ced-3、ced-4在細胞凋亡上是缺一不可的,而ced-9功用是抑制細胞凋亡,其抑制方法就是把ced-4接在一起附在粒線體膜上,而不讓ced-4是出去活化ced-3(如下圖),而要執行細胞凋亡的關鍵基因就是egl-1,egl-1會與ced-9結合,取代ced-4的位置,所以ced-4就會被釋出,進而一連串反應產生細胞凋亡(如下圖)。
訊息傳遞路徑
當死亡訊息(Ligand)傳遞到細胞膜表面的時候,此Ligand會與receptor做結合,而與ligand做結合後的receptor,其會透過膜內面的DD(deathdomain)來徵招(recruit)一個同樣具有DD的adaptor,這個adaptor的功用即是作為一個仲介者,它利用它上面一個DED(deatheffectordomain)來繼續往下徵招另一個proteinProcaspase8(上面同樣具有DED),而此Procaspase8即會切割並形成Caspase8,再來
Caspase8就會繼續往下活化,直到Caspase3被活化,遂行細胞凋亡。
但是,對於一些細胞來說,其透過訊息傳遞路徑所活化的Caspase3並不足夠,所以此時會透過一個Bid的protein,來將訊息傳往粒線體,一但Bid到達粒線體後,就會與粒線體上的Bcl-2結合,此時將會引發粒線體膜通透性增加,釋放出可誘發細胞凋亡的物質,所以,經由訊息傳遞路徑所誘導的細胞凋亡亦可經過粒線體加以放大。
粒線體路徑
由於此路徑是由細胞內部的粒線體所發動的,所以又叫intrinsicpathway,而誘使粒線體發動細胞凋亡的因子有許多種,如DNAdamage、毒物、氧化劑等等原因,一但粒線體接收到死亡訊息的時候,它的膜通透性就會增加,釋放出諸多物質可直接或間接地引發細胞凋亡;
其中就間接引發細胞凋亡的部份,粒線體會釋放出cytochromec,而此細胞色素可與Apaf-1此factor還有dATP結合成一個「小複合體」,而每七個這種小複合體會組成一個大的複合體,名為Apoptosome,這個Apoptosome可以活化Procaspase9此protein,使之形成Caspase9,而此protein會繼續往下活化細胞凋亡的執行者Caspase3,而這種一步一步往下傳的反應,稱作Caspasecascade,相當為一瀑布效應(同樣的,由細胞膜受體的訊息傳遞路徑來說,也會有Caspasecascade)。
圖示(Apoptosom)
粒線體除了釋放出cytochromec外,還可以釋放出其他跟apoptosis有關的物質,有可以直接引發細胞凋亡,如EndoG,就是一種DNase;
而Smac可以將Caspase3的inhibitXIAP移除,這是間接引發apoptosis的部份。
圖示(粒線體釋放之物質)
其中粒線體釋放出各種物質,是透過粒線體上的通道被打開;
而這個打開的機制是因和一類的protein有關,此類蛋白被稱為Bcl-2family,這個family主要的功能就是在調控粒線體上的膜通道,又此類蛋白可再分為三個子family,其中之一為促進apoptosis的protein,如Bcl-2;
另外兩個sub-families則是相反的功能,換句話說,是做為拮抗的作用,也就是anti-apoptosis,分別如Bax、Bid。
由於促進粒線體膜通道產生的假說非常多,若以其中之一的假說為例,本來可形成通道的Bax在平時,會與Bcl-2做結合而分開,也就是無法形成通道,一旦細胞接受到死亡訊號的時候Bid(屬於BH3onlyprotein,為一anti-apoptosis的Bcl-2sub-family)會與Bcl-2做結合,此結合將Bcl-2從Bax奪走,如此Bax將不再受Bcl-2所抑制,進而能形成膜通道。
圖示(粒線體膜通道形成機制)
和細胞凋亡有關的疾病
細胞凋亡的機制對人體是不可會缺的,所以當細胞凋亡出現了差錯,不論是過多(inordinate),無法控制,或是細胞凋亡的機制不足(insufficient),皆會對人體產生影響,造成疾病的產生。
Alzheimer’sdisease(因為inordinateapoptosis):
阿茲海默氏症是因為腦內的神經細胞不正常的萎縮,因此會造成人的記憶力喪失,所以又稱老人癡呆症,他主要是因為p53的作用使神經細胞凋。
機制;
機制和APP有關,APP是在神經細胞膜上的貫膜蛋白,正常時會跟G0蛋白還有PAK3結合,因此是穩定狀態,且此時細胞處G0期。
但在有家族性阿茲海默是正的人體內,APP不會跟G0結合,故他的切割酶便切割APP,切成A-beta及C99單元,接著C99再被切割為C57跟C31,這兩者皆會使細胞進入細胞周期的S期,於是遇到CHECHPOINT,再由P53的累積,導致神經細胞凋亡。
SLE(insufficientapoptosis)
即紅斑性狼瘡,他是因為細胞凋亡不完全,造成免疫機制不正常啟動,使皮膚出現紅斑性的狼瘡發炎反應。
在SLE的人體內,可能因家族遺傳,UV照射,或一些環境因子影響,使巨噬細胞上辨識PS的受體的DNA受損,故已細胞凋亡的細胞無法被吞噬,接下來變細胞壞死,於是是放危險訊號,或是巨噬細胞吞是這壞死細胞,表現抗原,這兩者皆會引起免疫機制,故發生發炎反應,表現在皮膚,變是紅斑性狼瘡。
參考資料
•科學月刊第三十三卷第十二期
•Apoptosis介紹(fromWikipedia)
•Replicationcycle介紹(fromWikipedia)
•細胞凋亡的分子機制
•Apoptosis簡介和相關圖片
•科學人雜誌相關報導
•FasLigand相關介紹
•ApoptosisandCancer比較報告
•細胞凋亡簡介及相關表格整理


- 配套讲稿:
如PPT文件的首页显示word图标,表示该PPT已包含配套word讲稿。双击word图标可打开word文档。
- 特殊限制:
部分文档作品中含有的国旗、国徽等图片,仅作为作品整体效果示例展示,禁止商用。设计者仅对作品中独创性部分享有著作权。
- 关 键 词:
- 大一 普通 生物 报告
 冰豆网所有资源均是用户自行上传分享,仅供网友学习交流,未经上传用户书面授权,请勿作他用。
冰豆网所有资源均是用户自行上传分享,仅供网友学习交流,未经上传用户书面授权,请勿作他用。
 转基因粮食的危害资料摘编Word下载.docx
转基因粮食的危害资料摘编Word下载.docx
-
高中英语词组大全Word文档下载推荐.docx
-
卫计局年工作总结及新年工作计划Word格式.docx
-
贵州省煤矿安全管理人员安全资格证A考试概况Word格式.docx
-
系统集成项目招标文件Word文件下载.docx
-
消防设计技术审查的要点Word文档格式.docx
-
第三章 习题课 带电粒子在磁场或复合场中的运动Word格式.docx
-
湖南岳阳中考英语模拟卷含答案Word文档格式.docx
-
电子商务考试题总汇打印版打印打印Word下载.docx
-
选调生考试备考言语理解与表达真题Word文档格式.docx
-
高考物理实验题专练 专练15Word文档格式.docx
-
加装奥迪A4L蓝牙电话功能Word文档下载推荐.docx
-
学年下学期好教育高三月考仿真卷A卷 语文 学生版后附详解Word文档下载推荐.docx
-
净化生产车间工程一般施工技术施工方案Word文档格式.docx
-
内蒙古呼和浩特市第六中学学年高一政治下学期期末考试试题Word下载.docx
-
证券行业客户经理电话营销技巧与实例Word文档下载推荐.docx
-
叶芝 苇间风文档格式.docx
-
最新中美贸易摩擦的原因及解决对策1论文Word文件下载.docx
-
意义的近义词Word格式文档下载.docx
-
上海市中考英语试题S.docx
-
专题12观点论证类设问.docx
-
附加安心重疾条款.docx
-
设计变更管理办法修改意见稿FINAL汇编.docx
-
毕业赠言毕业致词精选多篇.docx
-
银行新员工代表发言稿精选多篇.docx
-
北京市朝阳区届高三第一学期期末语文试题Word版含答案.docx
-
HL线切割使用说明书模板.docx
-
车工实训周记.docx
-
USBHID键盘扫描码.docx
-
Apmpoqu4调研报告.docx
-
最熟悉的陌生人作文八篇.docx
-
被动语态综合讲解.docx
-
幼儿教师年度思想工作总结4篇.docx
-
祝寿词集锦.docx
-
艺术导论结课论文1000字.docx
-
英语小技巧.docx
-
整理一年级课外阅读100篇.docx
-
致新人的一封信.docx
-
中华人民共和国海关总署商品归类决定世界海关组织至商品归类决定.docx
-
最新初中数学教师培训心得体会.docx
-
新课程背景下转变教学方式的研究.docx
-
珠宝营销学.docx
-
桩基检测方案低应变超声波钻芯及高应变法 2.docx
-
中国证券业协会反洗钱工作指引.docx
-
最新小学五级语文课外短文阅读训练教学内容.docx
-
中西药配伍禁忌表.docx
-
转载 《患者常见问题答疑》.docx
-
易燃易爆场所防雷装置及防静电接地装置检测技术规范.docx
-
中石油职称英语等级考试资料大全.docx
-
总机话务员先进事迹共8篇.docx
-
钻孔灌注桩技术规范施工规范监理细则.docx
