 第六章种子萌生文档格式.docx
第六章种子萌生文档格式.docx
- 文档编号:13534994
- 上传时间:2022-10-11
- 格式:DOCX
- 页数:16
- 大小:483.01KB
第六章种子萌生文档格式.docx
《第六章种子萌生文档格式.docx》由会员分享,可在线阅读,更多相关《第六章种子萌生文档格式.docx(16页珍藏版)》请在冰豆网上搜索。
种子吸胀作用并非活细胞的一种生理现象,而是胶体吸水体积膨大的物理作用。
由于种子的化学组成主若是亲水胶体,当种子生活力丧失以后,这些胶体的性质可不能相应发生显著转变。
因此不论是活种子或是死种子均能吸胀。
在一些情形下,活种子也会因种皮不透水而不能吸水膨胀。
种子吸胀能力的强弱,要紧决定于种子的化学成份。
高蛋白种子的吸胀能力远比高淀粉含量的种子为强,如豆类作物种子的吸水量大致接近或超过种子本身的干重,而淀粉种子吸水一样约占种子干重的1/2。
至于油料种子那么要紧决定于含油量的多少,在其它化学成份相似时,油分愈多,吸水力愈弱。
有些植物种子的外表有一薄层胶质,能使种子吸取大量水分,以供给内部生理的需要,亚麻种子确实是一例。
种子吸胀时,由于所有细胞体积增大,对种皮产生专门大的膨压,可能致使种皮破裂。
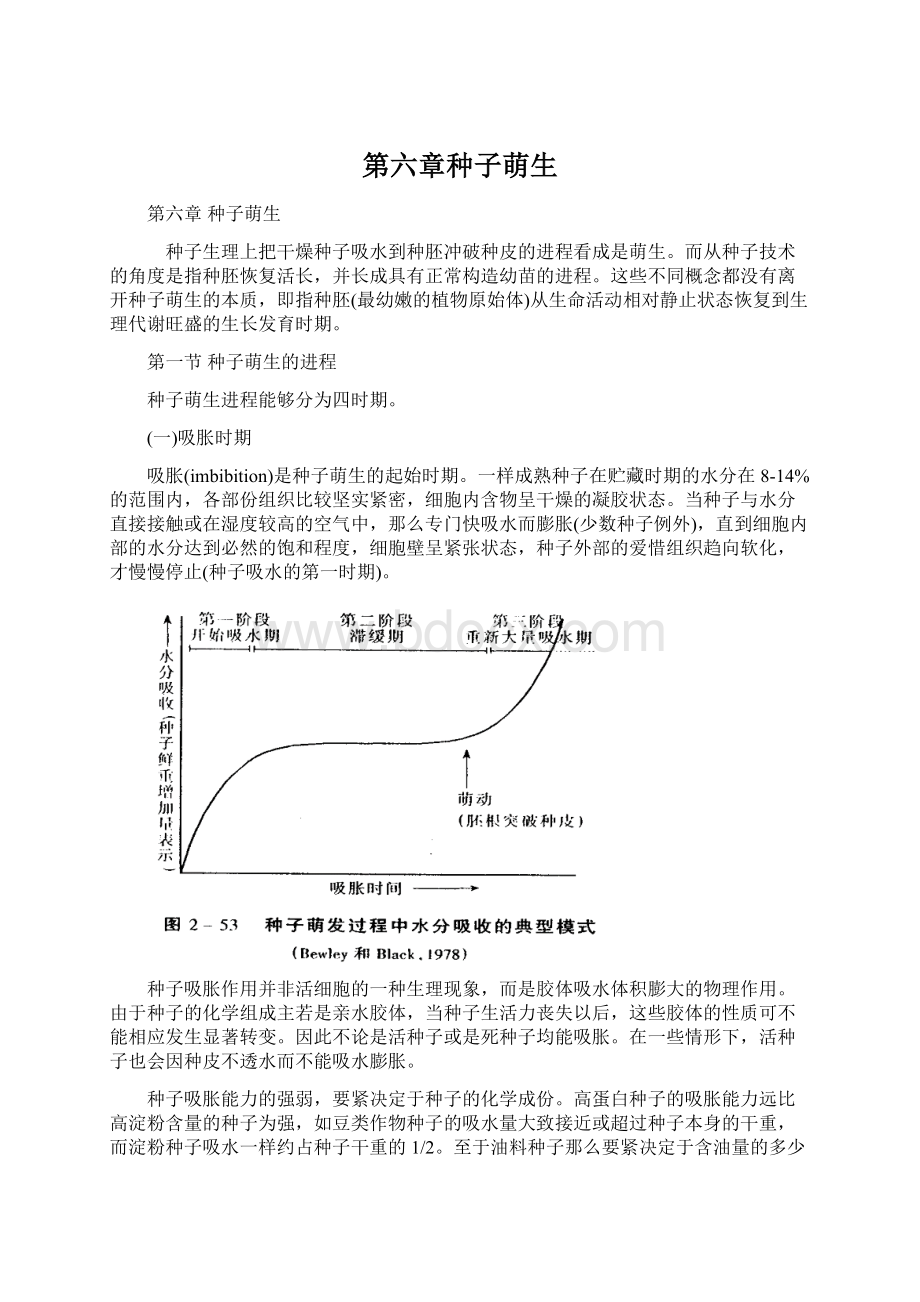
种子吸水达到必然量时(图2-53的第一时期终止)吸胀的体积与气干状态的体积之比,称为吸胀率。
一样淀粉种子的吸胀率是130-140%,而豆类种子的吸胀率达200%左右。
伴随吸胀进程,种胚活细胞内部的蛋白质、酶等大分子和细胞器等陆续发生水合活化。
例如当种子水分达16-18%,线粒体活性快速上升,光敏素活化。
在25℃下,玉米吸胀的前8h,菜豆吸胀最初10-16h,呼吸强度持续增加。
而死种子尽管能吸胀,但已经丧失了活化和修复能力。
有的死种子,由于蛋白质变性,原生质膜的透性提高,胶体的亲水性和保水能力降低,而使死种子的水分平稳大大改变,所吸收的水分充满这种死种子的细胞间隙和胚与胚乳的空间,呈现典型的水肿状态。
当死种子显现水肿现象,体积的膨大超级明显。
(二)萌动时期
萌动(protrusion)是种子萌生的第二时期。
种子在最初吸胀的基础上,吸水一样要停滞数小时或数天。
吸水尽管临时停滞,但种子内部的代谢开始增强,转入一个新的生理状态。
这一时期,在生物大分子、细胞器活化和修复基础上,种胚细胞恢复活长。
当种胚细胞体积扩大伸展到必然程度,胚根尖端就冲破种皮外伸,这一现象称为种子萌动。
种子萌动在农业生产上俗称为“露白”,说明胚部组织从种皮裂痕中开始显现出来的状况。
而种子生理学家适应上把萌动的到来看成是种子萌生的完成。
绝大多数植物的种子萌动时,第一冲破种皮的部份是胚根,因为胚根的尖端正对着种孔(发芽口),当种子吸胀时,水分从种孔进入种子,胚部优先取得水分,而且最先开始活动。
种子萌动时,胚的生长随水分供给情形而不同:
当水分较少时,那么胚根先出;
而当水分过量时,那么胚芽先出。
这是因为胚芽对缺氧的反映比胚根灵敏性差。
种子一开始萌动,其生理状态与休眠期间相较,即起了显著的转变。
胚部细胞的代谢性能趋向旺盛,而对外界环境条件的反映超级灵敏。
如碰到环境条件的急剧转变或各类理化因素的刺激,就可能引发生长发育失常或活力下降,严峻的会致使死亡。
在适当的范围内,给予或改变某些条件,会对整个萌生进程及幼苗的生长发育产生必然的效应。
(三)发芽时期
种子萌动以后,种胚细胞开始或加速割裂和分化,生长速度显著加速,当胚根、胚芽伸出种皮并发育到必然程度,就称为发芽(germination)。
我国和国际种子查验规程对发芽概念是当种子发育长成具有正常要紧构造的幼苗才称为发芽。
种子处于这一时期,种胚的新陈代谢作用极为旺盛,呼吸强度达最高限度,会产生大量的能量和代谢产物。
若是氧气供给不足,易引发缺氧呼吸,放出乙醇等有害物质,使种胚窒息麻痹以致中毒死亡。
农作物种子如催芽不妥,或播后受到不良条件的阻碍,常会发生这种情形。
例如大豆、花生及棉花等大粒种子,在播种后由于土质粘重、密度过大或覆土过深、雨后表土板结,种子萌动会因氧供给不足,呼吸受阻,生长停滞,幼苗无力顶出土面,而发生烂种和缺苗断垄等现象。
能量的利用-----种子发芽进程中所放出的能量是较多的,其中一部份热量散失到周围土壤中;
另一部份成为幼苗顶土和幼根入土的动力。
健壮的农作物种子出苗快而整齐,消瘦的种子营养物质少,发芽时可利用的能量不足,即便播种深度适应,亦常常无力顶出而死亡;
有时虽能出土,但因活力很弱,经不起恶劣条件的侵袭,一样容易引发死苗。
(四)幼苗的形态建成(seedlingestablishment)?
v种子发芽后依照其子叶出土的状况,可分成两种类型的幼苗。
一、子叶出土型
双子叶的子叶出土型(epigealgermination)植物在种子发芽时,其下胚轴显著伸长,初期弯成拱形,顶出土面后在光照诱导下,生长素散布相应转变,使下胚轴慢慢伸直,生长的胚与种皮(有些种子连带小部份残余胚乳)离开,子叶迅速展开,见光后慢慢转绿,开始营光合作用,以后从两子叶间的胚芽长出真叶和主茎。
单子叶植物中只有少数属子叶出土型,如葱蒜类等,而90%的双子叶植物幼苗属这种类型,常见的作物有棉花、油菜、大豆、黄麻、烟草、蓖麻、向日葵和瓜类等。
这种植物幼苗下胚轴的长度和生长快慢与出苗率常有紧密关系。
子叶出土型幼苗的优势是幼苗出土时顶芽包被在子叶中受到爱惜,子叶出土后能进行光合作用,继续为生长提供能量,象大豆的子叶能进行数日的光合作用,而棉花、萝卜等子叶能维持数周的光合功能。
某些植物的子叶与后期生育有关,如棉花的子叶受到损害时,以后会减少结铃数,乃至完全不结铃;
丝瓜的子叶受伤后,对开花期子房的发育会产生抑制作用,因此在作物移植或间苗操作进程中,就注意爱惜子叶的完整,幸免机械损伤。
二、子叶留土型
双子叶的子叶留土型(hypogealgermination)植物在种子发芽时,上胚轴伸长而出土,随即长出真叶而成幼苗,子叶仍留在土中与种皮不离开,直至内部贮藏养料消耗殆尽,才萎缩或解体。
大部份单子叶植物种子,如禾谷类,小部份双子叶植物种子,如蚕豆、豌豆、茶叶属于这一类型,后者的子叶一样较肥厚。
这种留土型的种子发芽时,穿土力较强,即便在粘重的土壤中,一样也较少发觉闭孔现象。
因此,播种时可较出土型的略深,尤其在干旱地域,更属必要。
禾谷类种子幼苗出土的部份事实上是“子弹型”的胚芽鞘,胚芽鞘出土后在光照下开裂,内部的真叶才慢慢伸出,进行光合作用。
如没有完整胚芽鞘的爱惜作用,幼苗出土将受到阻碍。
另外,由于留土幼苗的营养贮藏组织和部份侧芽仍保留在土中,因此一旦土壤上面的幼苗部份受到昆虫、低温等的损害,仍有可能从头从土中长出幼苗。
第二节种子萌生的生理生化进程
(一)细胞的活化和修复
在成熟的干种子细胞内部预存着一系列生命代谢和与合成有关的生化系统,在种子萌生的最初时期,细胞吸水后当即开始修复和活化活动。
活化和修复在吸水的第一、二两个时期进行。
细胞膜修复----正常的细胞膜中,磷脂和膜蛋白的排列整齐,结构完整。
当种子成熟和干燥进程中,由于种子脱水,磷脂的排列发生转向,膜的持续界面再也不能维持,膜成为不完整状态。
以致种子吸水以后,细胞膜失去其正常的功能,无法避免溶质从细胞内渗漏出去。
吸胀一按时刻以后,种子内修补细胞膜的进程完成,膜就恢复了正常的功能,溶质的渗出就取得了阻止。
现还发觉,吸胀细胞新合成磷脂分子,在高水分下磷脂和膜蛋白分子在细胞膜上排列趋向完整。
随着种子吸胀的进行,线粒体内膜的某些缺损部份从头合成,恢复完整,电子转移酶类被合成或活化并嵌入膜中,结果氧化磷酸化的效率慢慢恢复正常。
DNA修复------DNA分子损伤的修复由DNA内切酶、DNA多聚酶和DNA连接酶来完成。
修复的一样方式是第一由内切酶切去受到损伤的片段,接着由多聚酶从头合成相应片段,再由连接酶连接到相应DNA分子上。
而一样的DNA分子裂口可由连接酶作直接的接合。
干种子中缺损的RNA分子一样被分解,而由新合成的完整RNA分子所取代。
活化和修复能力,除受环境条件的阻碍外,还与种子的活力有紧密关系。
低活力的种子活化迟缓,修复困难。
因为低活力的种子不仅修复能力降低,而且损伤的程度比高活力的种子大得多,活力降低到必然水平,就无法修复,种子也就失去萌生能力。
(二)种胚的生长和合成代谢
种子萌生最初的生长在种胚细胞内要紧表此刻活化和修复基础上细胞器和膜系统的合成增殖。
修复时原有线粒体的部份膜被合成,呼吸酶数量增加,呼吸效率大大提高;
而接着细胞中新线粒体形成,数量进一步增加。
同时,细胞的内膜系统——内质网和高尔基体也大量增殖。
高尔基体运输多糖到细胞壁作为合成原料;
内质网能够产生小液泡,小液泡的吸水胀大和液泡间的融合,使胚根细胞体积增大。
在许多情形下,胚根细胞的伸长扩大,就可直接致使种子萌动。
种胚细胞具有很强的生长和合成能力。
以小麦种子为例,吸胀30min即利用种子预存的RNA合成蛋白质;
新RNA分子的合成在吸胀后3h开始,第一合成的种类是mRNA。
在必然量的新RNA积存的基础上,小麦种子中DNA的合成于吸胀的第15h开始,在DNA复制后数小时,种胚细胞进行有丝割裂。
(三)贮藏物质的分解和利用
种子内部存在丰硕的营养物质,在发芽进程中慢慢地被分解和利用。
一方面在呼吸进程中转化为能量,用于生长和合成;
另一方面通过代谢转化成新细胞组成的成份。
在种子吸胀萌动时期,生长先动用胚部或胚中轴(embryoaxis)的可溶性糖、氨基酸和仅有少量的贮藏蛋白。
贮藏组织(胚乳或子叶)中贮藏物质的分解需在种子萌动以后。
淀粉、蛋白质和脂肪等大分子第一被水解成可溶性的小分子,然后输送到胚的生长部位被继续分解和利用(图2-55)。
要紧贮藏物质分解利用的方式如下:
一、淀粉
90%的淀粉水解成葡萄糖要紧由淀粉水解酶所催化,α-淀粉酶的产生与GA的诱导有关,而β-淀粉酶要紧预存在胚乳中。
禾谷类种子的盾片在萌生中具有分泌和消化吸收的功能,在淀粉的分解中也起重要作用(图2-56)。
二、蛋白质
种子蛋白质的分解是分步进行的。
第一步是贮藏蛋白可溶化,非水溶性的贮藏蛋白不易直接被分解成氨基酸,第一被部份水解形成水溶性的分子量较小的蛋白质;
第二步是可溶性蛋白完全氨基酸化,可溶性蛋白被肽链水解酶(包括肽链内切酶、羧肽酶、氨肽酶)水解成氨基酸。
这种蛋白质水解的时期性在双子叶种子中表现得专门明显。
禾谷类种子蛋白质的分解要紧发生在三个部位:
(1)胚乳淀粉层
(2)糊粉层
(3)胚中轴和盾片
贮藏蛋白质分解成氨基酸从头组成蛋白质的进程中,很多氨基酸未被直接利用而进行转化。
这些氨基酸通过氧化脱氨作用,进一步分解为游离氨及不含氮化合物。
因此很容易发觉游离氨的存在。
这种游离氨如积存过量,就会使植物细胞中毒。
在一样情形下,游离氨的存在量很少。
细胞中含有足够的糖类时,游离氨直接进入氨基化反映,和糖类所衍生的酮酸形成新氨基酸,再从头合成蛋白质。
3、脂肪
存在于细胞质脂质体中的脂肪第一被脂肪水解酶水解成甘油和脂肪酸。
脂肪酸在乙醛酸体中进行β-氧化,生成乙酰CoA进入到乙醛酸循环。
乙醛酸循环产生的琥珀酸转移到线粒体中通过三羧酸循环形成草酰乙酸,再通过糖酵解的逆转转化为蔗糖,输送到生长部位。
甘油能在细胞质中迅速磷酸化,随后氧化为磷酸丙糖,在醛缩酶的作用下缩合成六碳糖


- 配套讲稿:
如PPT文件的首页显示word图标,表示该PPT已包含配套word讲稿。双击word图标可打开word文档。
- 特殊限制:
部分文档作品中含有的国旗、国徽等图片,仅作为作品整体效果示例展示,禁止商用。设计者仅对作品中独创性部分享有著作权。
- 关 键 词:
- 第六 种子 萌生
 冰豆网所有资源均是用户自行上传分享,仅供网友学习交流,未经上传用户书面授权,请勿作他用。
冰豆网所有资源均是用户自行上传分享,仅供网友学习交流,未经上传用户书面授权,请勿作他用。
 《稻草人》精彩读书笔记精选多篇.docx
《稻草人》精彩读书笔记精选多篇.docx
-
《鸿门宴》原文及注解doc资料.docx
-
《张石匠拜师》教学设计.docx
-
1万亩滴灌鲜食葡萄建设项目可行性研究报告.docx
-
《长方体和正方体整理与复习课》说课稿.docx
-
7A文铁路隧道工程施工承包合同中英文.docx
-
《纪念刘和珍君》学案1.docx
-
《我们之间》读后感范文6篇.docx
-
BRW200315型乳化液泵组随机图册.docx
-
STM32实现万年历.docx
-
《保险学》考试复习资料要点.docx
-
《讲究卫生预防疾病》国旗下讲话.docx
-
《三国演义》读书笔记摘抄赏析好词好句读后感悟范文.docx
-
《路基路面工程》每章练习题.docx
-
《谛听教育的春天》读书笔记.docx
-
17春学期清考《市政学》在线作业1.docx
-
201X商品房现房买卖合同范本word范文 10页.docx
-
《欧姆定律》课件.docx
-
04上海虹口区高三历史二模课案.docx
-
7A版南大考研真题整理版.docx
-
《蝙蝠和雷达》教学设计.docx
-
2450m3高炉及225000kva矿热炉项目节能论证评估报告.docx
-
《商业银行管理学》课后习题答案.docx
-
《论语十二章》练习答案.docx
-
3年高考2年模拟1年原创系列小说阅读教案.docx
-
《小学数学生活化的研究》开题报告.docx
-
9A文汽车换挡机构设计指南.docx
-
《柴油机》课程教学设计.docx
-
《股票作手回忆录》读后感精选多篇.docx
-
《新闻纵横》生存类民生新闻的分析报告.docx
-
《科学探索者》丛书参考答案天文学.docx
-
《网络聊天那点事》5人年会晚会搞笑小品剧本台词.docx
-
成都市电子政务云平台项目建议书.docx
-
部编七年级历史下册专项试题.docx
-
初二家长会数学教师发言稿多篇.docx
-
财政财务收支审计报告模板.docx
-
成品化粪池施工方案.docx
-
初一学生旷课检讨书doc.docx
-
春春体艺2+1计划.docx
-
初中政治教师年度工作总结.docx
-
初级育婴师年考试试题加标准答案.docx
-
超星网课领导学答案.docx
-
初中生课外诗词诵读与鉴赏指导研究报告.docx
-
初一生物复习资料整合.docx
-
电工电子综合实验.docx
-
创建优质中学实现跨越发展田心中学中学创优汇报资料.docx
-
电子商务信安全习题.docx
-
初中文言文虚词集锦.docx
-
部编版七年级语文上册第六单元测试题.docx
-
彩板房的结构受力特点及节点设计正文.docx
-
餐饮服务食品安全系统管理系统人员必备知识参考题库.docx
